


ISSN: 0973-7510
E-ISSN: 2581-690X
Multidrug-resistant (MDR) bacteria have posed a major clinical challenge, as many currently available antibiotics have lost their effectiveness. The development of new or improved antimicrobial formulations are required to address this problem. Collected nosocomial multidrug-resistant bacteria were tested for their susceptibility to various aminoglycosides to determine the minimum inhibitory concentrations (MICs). The most potent isolate producing a modifying enzyme inhibitor was identified. The conditions for maximal inhibitory activity were optimized, and the inhibitory protein was subsequently purified and combined with antibiotics to formulate an active preparation against resistant bacteria. The cytotoxicity of the inhibitor was evaluated using a human skin cell line. Streptococcus pneumoniae (B25) exhibited resistance to amikacin, tobramycin, gentamicin, streptomycin, neomycin, kanamycin, and paromomycin, with MICs of 500, 440, 320, 170, 110, 200, and 140 µg/mL, respectively. Similarly, Staphylococcus aureus (B75) was resistant to the same antibiotics, showing MICs of 80, 140, 200, 470, 440, 500, and 410 µg/mL, respectively. Pseudomonas aeruginosa (B80) was the most effective isolate, exhibiting strong inhibitory activity against both S. pneumoniae (B25) and S. aureus (B75). The inhibitory protein was precipitated at 70% ammonium sulfate saturation, purified by ion-exchange and gel-filtration chromatography, and appeared as a single 20 kDa band on SDS-PAGE. Paromomycin (140 or 410 µg/mL) and streptomycin (170 or 470 µg/mL), when combined with a modifying enzyme inhibitor (61.44 µg/mL), showed a significant increase in antibacterial activity against S. pneumoniae (B25) and S. aureus (B75). The inhibitory protein exhibited less than 5% cytotoxicity toward the human skin fibroblast (HSF) cell line, maintaining approximately 99% cell viability.
Multidrug-resistance, Modifying Enzymes, Antagonism, Aminoglycosides, Optimization
Aminoglycosides are a potent class of antibiotics widely used to treat severe infections; however, the emergence of bacterial resistance through various mechanisms has become a significant clinical concern. This resistance problem can, to some extent, be mitigated by certain bacterial strains that counteract aminoglycoside-resistant bacteria through antagonistic interactions. Multidrug-resistance (MDR) was first recognized approximately 50 years ago and remains a critical global health issue, particularly in developing countries, where antibiotic misuse and other medical factors accelerate its spread. Aminoglycosides constitute a broad-spectrum group of antibiotics that serve as first-line treatments for several serious infections, including urinary tract, gastrointestinal, respiratory infections, and sepsis. Nonetheless, some Gram-positive and Gram-negative bacterial strains exhibit high-level resistance to aminoglycosides, complicating therapeutic management. Among Gram-positive pathogens, methicillin-resistant Staphylococcus aureus (MRSA) represents one of the most common MDR bacteria, displaying resistance to β-lactams, aminoglycosides, and macrolides.1 MRSA can inactivate aminoglycosides through the secretion of modifying enzymes such as nucleotidyl transferases, encoded by plasmid- or chromosome-borne genes including aac(6’), aph(2″), aph(3’)-IIIa, and ant(4’)-Ia.2
According to repeated reports from the World Health Organization (WHO) since 1990, S. aureus and Streptococcus pneumoniae remain the most prevalent nosocomial pathogens, particularly in hospital nurseries and intensive care units. In developing regions, multidrug-resistant S. pneumoniae poses a major clinical threat due to its virulence and its ability to evade treatment with conventional antibiotics. Resistance in S. pneumoniae involves multiple mechanisms, including efflux pumps, enzymatic modification, and DNA methylation. Epidemiological data indicate that resistant pneumococcal strains disseminate rapidly, primarily through genetic transfer. Although newer antibiotics such as lefamulin and gepotidacin have been introduced as first-line agents against MDR bacteria, the continued development of novel or improved antibiotics remains essential to address this escalating threat.3 The spread of multidrug-resistance (MDR) among humans is primarily attributed to the overuse and misuse of antibiotics, particularly through direct or indirect contact with poultry infected by multidrug-resistant pathogenic bacteria.4 Two main factors contribute to the development of MDR: physiological stress caused by infections and the inappropriate use of antibiotics.5 To address the declining efficacy of conventional antibiotics, the development and application of alternative antimicrobial agents are essential. Several promising alternatives have been identified, including medium-chain fatty acids,6 plant extracts,7-11 antimicrobial peptides,12 as well as prebiotics and probiotics.13-15
Many serious human diseases, including arthritis, sepsis, pneumonia, and even death are associated with Gram-positive bacteria, particularly S. aureus and S. pneumoniae. Gram-negative pathogens, such as Escherichia coli and Enterococcus spp., are also significant contributors to infectious diseases.16 S. aureus is responsible for a wide range of conditions, from dermatitis to multi-organ failure, and it is also the most prevalent bacterial pathogen in animals, especially poultry.17 Both MRSA and methicillin-resistant S. pseudintermedius (MRSP) represent major public and veterinary health threats.18 Globally, S. aureus and S. pneumoniae are among the most prominent Gram-positive bacteria exhibiting resistance to multiple antibiotic classes, including aminoglycosides, fluoroquinolones, macrolides, and sulfonamides. Reports from various countries, including the USA,19 Germany,20 Poland,21 Hong Kong,22 and South Korea23 highlight the global scale of this resistance. The Staphylococcus genus comprises approximately 104 species, classified as either commensal or pathogenic. Staphylococci are opportunistic organisms commonly found as commensals in water, soil, and surfaces, or as part of the normal flora of the skin, digestive, and respiratory tracts of humans and animals. However, they can become pathogenic under conditions such as wounds or tissue injury.24 Similarly, S. pneumoniae is a major human pathogen responsible for diseases ranging from otitis media to severe systemic infections. Pneumococci are also highly opportunistic in animals, frequently causing secondary infections in waterfowl that often result in mortality, particularly during the first three weeks of life.25
Aminoglycoside-modifying enzyme inhibitors have emerged as a promising approach in overcoming bacterial resistance by directly targeting the catalytic activity of modifying enzymes. These inhibitors function through diverse mechanisms, including competitive binding to enzyme active sites, allosteric modulation to induce conformational changes, or cofactor mimicry to disrupt enzymatic reactions. Various classes of inhibitors have been explored, ranging from natural products such as flavonoids and polyphenols, which exhibit selective inhibitory activity, to synthetic small molecules designed as bisubstrate analogues that simultaneously mimic substrates and cofactors. Peptide-based inhibitors are also under investigation for their ability to selectively obstruct enzyme activity, while dual-function compounds are being developed to combine direct antimicrobial effects with enzymatic inhibition.25
Despite their potential, the development of effective inhibitors faces several challenges. Achieving specificity across diverse enzyme isoforms remains difficult, and potential off-target effects on human enzymes raise safety concerns. Furthermore, pharmacokinetic limitations, including stability and bioavailability, can restrict therapeutic efficacy. Nonetheless, preclinical studies have demonstrated that these inhibitors can restore enzymatic susceptibility in resistant bacterial strains, highlighting their potential as adjunctive agents. Ongoing research focuses on optimizing potency, selectivity, and delivery, aiming to translate these inhibitors into clinically viable options for combating resistance.25
This study aims to control aminoglycoside-resistant bacteria by utilizing an inhibitory protein or modifying enzyme produced by another microorganism through a biological phenomenon known as antagonism. This approach aligns with the broader objectives of antimicrobial resistance research, focusing on the biological control of drug-resistant pathogens. Additionally, the study seeks to optimize the production and purification of the inhibitory protein to achieve synergistic effects with antibiotics, thereby enhancing the inhibition of resistant bacterial strains.
Aminoglycosides
The aminoglycosides used in this study: amikacin, streptomycin, tobramycin, gentamicin, neomycin, kanamycin, and paromomycin were purchased from Sigma (USA).
Collection of pathogenic bacteria
Pathogenic bacterial isolates were collected from King Khalid and Al-Salam Hospitals during January-February 2025 using sterile swabs. Each swab was immersed in nutrient broth under aseptic conditions for 30 sec and incubated at 37 °C for 48 hrs. A loopful of the resulting bacterial suspension was streaked onto nutrient agar plates and incubated at 37 °C for 24 hrs. Distinct colonies were selected using sterile loops, restreaked on fresh agar plates, and incubated again at 37 °C for 24 hrs to ensure purity. Homogeneous growth and uniform colony morphology were considered indicative of pure cultures, which were maintained on nutrient agar slants at 4 °C for up to one month and subcultured periodically to preserve viability.
Determination of MIC
The minimum inhibitory concentrations (MICs) of the tested antibiotics against the most resistant bacterial isolates were determined following the Clinical and Laboratory Standards Institute (CLSI) guidelines.26,27 Prior to MIC testing, bacterial isolates were cultured overnight in 3 mL of Mueller–Hinton broth (MHB) and incubated at 37 °C for 24 hrs. MIC assays were performed using sterile 96-well microtiter plates (VWR International, LLC, Debrecen, Hungary). All wells, except those in the first column, were prefilled with 90 µL of MHB. Serial twofold dilutions of antibiotic stock solutions prepared in MHB were made, starting from 530 µg/mL and covering a concentration range of 530-20 µg/mL. Subsequently, 180 µL of each antibiotic dilution was added to the first column of the plate, followed by serial twofold dilutions across the remaining columns. A 10 µL aliquot of bacterial suspension adjusted to 0.5 McFarland standard (approximately 105 CFU/mL) was inoculated into each well from column 11 backward using a sterile tumbler (Thermo Fisher Scientific, Budapest, Hungary). Column 11 served as the positive control (containing only bacterial suspension and broth), while column 12 served as the negative control (containing only broth). Plates were incubated at 41 °C for 24 hrs. MIC values were determined using a Sensititre™ SWIN™ automated MIC reader (Thermo Fisher Scientific, Budapest, Hungary) with the VIZION™ system software (version 3.4, Thermo Fisher Scientific, 2024). S. aureus (ATCC 12600) and S. pneumoniae (ATCC 49619) were used as quality control reference strains.
Identification of bacterial isolates
Classic identification
Bacterial isolates were identified based on their morphological and biochemical characteristics according to Bergey’s Manual of Systematic Bacteriology (1989).
Genetic identification
Genomic DNA was extracted from bacterial isolates using the Epicentre MasterPure™ Complete DNA and RNA Extraction Kit (Mandel Scientific, Guelph, ON, Canada) following the manufacturer’s standard protocol. Molecular identification was performed by amplification and sequencing of the 16S rRNA gene using a thermocycler (Perkin-Elmer Model Cetus 480). PCR amplification was carried out with universal primers 27F (5′-AGA GTT TGA TCC TGG CTC AG-3′) and 1525R (5′-AAG GAG GTG ATC CAG CC-3′). The PCR program consisted of an initial denaturation at 94 °C for 5 min, followed by 35 cycles of denaturation at 94 °C for 60 sec, annealing at 55 °C for 60 sec, and extension at 72 °C for 90 sec, with a final extension at 72 °C for 5 min. PCR products were purified and sequenced using the BigDye™ Terminator Cycle Sequencing Kit (PE Applied Biosystems, USA) on an ABI 310 automated DNA sequencer (Applied Biosystems, USA). The obtained 16S rRNA gene sequences were compared with reference sequences in the GenBank database using the BLAST algorithm for species-level identification.28,29
Optimization of environmental conditions
Optimal environmental conditions, including incubation period, temperature, pH, stirring speed, aeration rate, and sources of carbon and nitrogen were optimized to maximize the production of the inhibitory protein produced by P. aeruginosa. The inhibitory activity was tested against amikacin, gentamicin, and tobramycin in combination with S. pneumoniae (B25), and against streptomycin, neomycin, kanamycin, and paromomycin in combination with S. aureus (B75), following the method described by Vanderzant and Don.30
Effect of incubation period
Nutrient broth (pH 7.2) was prepared and dispensed into 250 mL Erlenmeyer flasks (100 mL per flask), sterilized by autoclaving, and inoculated with 1 mL of P. aeruginosa culture (3 × 105 CFU/mL). The cultures were incubated at 37 °C with shaking at 120 rpm for 16, 20, 24, 28, 32, 36, 40, 44, and 48 hrs. Nutrient agar plates containing amikacin, gentamicin, and tobramycin were inoculated with S. pneumoniae (B25), while plates containing streptomycin, neomycin, kanamycin, and paromomycin were inoculated with S. aureus (B75). Wells were aseptically punched into the agar and filled with the P. aeruginosa culture corresponding to each incubation period. Plates were incubated at 37 °C for 24 hrs, and the diameter of the inhibition zones surrounding the wells was measured to assess the antibacterial activity of the inhibitory protein. All experiments were performed in triplicate.
Effect of incubation temperature
Nutrient broth (pH 7.2) was prepared and dispensed into 250 mL Erlenmeyer flasks (100 mL per flask), sterilized by autoclaving, and inoculated with 1 mL of P. aeruginosa culture (3 × 106 CFU/mL). The cultures were incubated with shaking at 120 rpm for 24 hrs at different temperatures (20, 25, 30, 35, 40, and 45 °C). Nutrient agar plates containing amikacin, gentamicin, and tobramycin were inoculated with S. pneumoniae (B25), while those containing streptomycin, neomycin, kanamycin, and paromomycin were inoculated with S. aureus (B75). Wells were aseptically drilled into the agar and filled with P. aeruginosa cultures grown at each temperature condition. Plates were then incubated at 37 °C for 24 hrs, and inhibition zones around the wells were measured to determine antibacterial activity. All experiments were performed in triplicate.
Effect of pH
Nutrient broth was prepared and dispensed into 250 mL Erlenmeyer flasks (100 mL per flask). The pH of each flask was adjusted to specific values (6.6, 6.8, 7.0, 7.2, 7.4, 7.6, 7.8, 8.0, and 8.2) using 0.5 M NaOH or 1 N HCl. The media were autoclaved, inoculated with 1 mL of P. aeruginosa culture (3 × 106 CFU/mL), and incubated at 37 °C with shaking at 120 rpm for 24 hrs. Nutrient agar plates containing amikacin, gentamicin, and tobramycin were inoculated with S. pneumoniae (B25), while those containing streptomycin, neomycin, kanamycin, and paromomycin were inoculated with S. aureus (B75). Wells were aseptically drilled into the agar and filled with P. aeruginosa cultures grown at each pH condition. The plates were incubated at 37 °C for 24 hrs, and inhibition zones around the wells were measured to assess antibacterial activity. All experiments were performed in triplicate.
Effect of stirring speed
Nutrient broth (pH 7.2) was prepared and dispensed into 250 mL Erlenmeyer flasks (100 mL per flask), sterilized by autoclaving, and inoculated with 1 mL of P. aeruginosa culture (3 × 106 CFU/mL). The cultures were incubated at 37 °C for 24 hrs with shaking at different speeds (100, 110, 120, 130, 140, 150, 160, 170, and 180 rpm). Nutrient agar plates containing amikacin, gentamicin, and tobramycin were inoculated with S. pneumoniae (B25), while those containing streptomycin, neomycin, kanamycin, and paromomycin were inoculated with S. aureus (B75). Wells were aseptically drilled into the agar and filled with P. aeruginosa cultures grown at each stirring speed. Plates were incubated at 37 °C for 24 hrs, and inhibition zones around the wells were measured to evaluate antibacterial activity. All experiments were performed in triplicate.
Effect of aeration
Nutrient broth (pH 7.2) was prepared and dispensed into 250 mL Erlenmeyer flasks containing different culture volumes (20, 40, 60, 80, 100, 120, 140, and 160 mL). The flasks were sterilized by autoclaving, inoculated with 1 mL of P. aeruginosa culture (3 × 106 CFU/mL), and incubated at 37 °C with shaking at 120 rpm for 24 hrs. Nutrient agar plates containing amikacin, gentamicin, and tobramycin were inoculated with S. pneumoniae (B25), while those containing streptomycin, neomycin, kanamycin, and paromomycin were inoculated with S. aureus (B75). Wells were aseptically drilled into the agar and filled with P. aeruginosa cultures grown under each aeration condition. Plates were incubated at 37 °C for 24 hrs, and inhibition zones were measured to assess antibacterial activity. All experiments were performed in triplicate.
Effect of carbon/nitrogen source
Nutrient broth (pH 7.2) was prepared by replacing yeast extract (as the primary carbon source) with different carbon sources: glucose, galactose, fructose, sucrose, mannitol, starch, and xylose and by replacing peptone (as the primary nitrogen source) with alternative nitrogen sources – KNO3, (NH4)2SO4, NH3, valine, histidine, and alanine. Each formulation was dispensed into 250 mL Erlenmeyer flasks (100 mL medium per flask). The flasks were autoclaved, inoculated with 1 mL of P. aeruginosa culture (3 × 106 CFU/mL), and incubated at 37 °C with shaking at 120 rpm for 24 hrs. Nutrient agar plates containing amikacin, gentamicin, and tobramycin were inoculated with S. pneumoniae (B25), while those containing streptomycin, neomycin, kanamycin, and paromomycin were inoculated with S. aureus (B75). Wells were aseptically drilled into the agar and filled with P. aeruginosa cultures grown with each specific carbon or nitrogen source. Plates were incubated at 37 °C for 24 hrs, and inhibition zones were measured to assess antibacterial activity. All experiments were performed in triplicate. The weights of alternative carbon or nitrogen sources were calculated using the following equations:
W = M × % of carbon/nitrogen in an alternative carbon/nitrogen source × 100
Where W is the weight of an alternative carbon/nitrogen source, M is the weight of original carbon/nitrogen source.
% of carbon/nitrogen in an alternative carbon/nitrogen source = Atomic mass / Molecular mass × 100
Preparation of cell free extract
Nutrient broth was inoculated with 1 mL of P. aeruginosa culture (3 × 106 CFU/mL) and incubated at 37 °C for 24 hrs. The bacterial culture was centrifuged at 5,000 × g for 10 min to separate the cells, and the resulting supernatant containing the extracellular inhibitory protein was collected. The protein was precipitated from the supernatant using saturated ammonium sulfate.
Ammonium sulfate precipitation of the inhibitory protein
The inhibitory protein was precipitated using a range of saturated ammonium sulfate solutions (30%-90%). Each fraction was allowed to stand for 2 hrs at 4 °C and then centrifuged at 5,000 × g for 10 min at 4 °C. The resulting pellet was dissolved in 10 mL of phosphate buffer (pH 7.5), heated at 70 °C for 10 min, cooled to room temperature, and centrifuged again at 5,000 × g for 10 min at 4 °C. The supernatant was collected for determination of the optimal ammonium sulfate saturation.
Protein quantification
An aliquot of the protein sample (1 mL) was transferred into a clean glass test tube and mixed with 5 mL of alkaline copper reagent. The mixture was left at room temperature for 15 min.31 Subsequently, 0.5 mL of diluted Folin Ciocalteu reagent was added, mixed gently to ensure homogeneity, and incubated at room temperature for 30 min. The absorbance of the resulting blue color was measured at 280 nm using a UV-Vis spectrophotometer. A blank was prepared using 5 mL of alkaline copper reagent mixed with 0.5 mL of diluted Folin Ciocalteu reagent.
Purification of the inhibitory protein
Ion-exchange chromatography
A glass chromatography column (2.5 × 50 cm) was carefully packed with 100 g of diethylaminoethyl cellulose (DEAE-cellulose) to avoid bubble formation. Prior to use, the resin was washed with distilled water, adjusted to pH 6.5 using 1 N HCl, and subsequently treated several times with 0.5 M NaOH until the wash solution became colorless. The resin was then thoroughly rinsed with distilled water to remove residual alkalinity and suspended in three volumes of phosphate buffer solution (pH 7.4). The suspension was allowed to settle, and the resulting precipitate free of fine particles was used to pack the column.
Gel filtration chromatography
A glass chromatography column (2.5 × 20 cm) was carefully packed with 10 g of Sephadex G-100. Prior to packing, the resin was swollen in 400 mL of phosphate buffer solution (pH 7.4), heated in a water bath for 6 h, then cooled to 50 °C before loading into the column.32 Fractions containing the inhibitory protein were pooled and dialyzed overnight against phosphate buffer solution (pH 7.4).
SDS-PAGE analysis
The purified inhibitory protein was mixed with 0.1% sodium dodecyl sulfate (SDS) at pH 8.8 containing 1 mM dithiothreitol (DTT), and electrophoresis was carried out on a 10% polyacrylamide gel using the SDS-PAGE technique.33 The gel and buffer systems were prepared according to the method of Laemmli.34 After electrophoresis, the gel was stained with Coomassie Brilliant Blue to visualize the protein bands.35
Determination of inhibitory activity
The inhibitory activity of antibiotics at their respective MIC values against S. pneumoniae (B25) and S. aureus (B75) was evaluated in the presence of varying concentrations of the purified inhibitory protein (0.015-245.76 µg/mL). Stock solutions of antibiotics and the inhibitory protein were prepared according to CLSI guidelines.26,27 Nutrient agar was prepared, cooled to 50 °C, and supplemented with defined concentrations of both the antibiotic and inhibitory protein. The mixture was thoroughly homogenized, poured into Petri dishes, and allowed to solidify. Plates were then inoculated with the resistant bacterial strains and incubated at 37 °C for 24 hrs.
Cytotoxicity of modifying enzyme inhibitor on human skin fibroblast (HSF) cell line
Cell culture and treatment
The HSF cell line, a normal human skin fibroblast line, was obtained from Catalent Biologics (Oxfordshire, UK). Cells were cultured in Dulbecco’s Modified Eagle’s Medium (DMEM) under a humidified atmosphere containing 5% (v/v) CO‚ at 37 °C. The culture medium was divided into five aliquots for treatment. The first aliquot served as the negative control and was supplemented with 10% heat-inactivated fetal bovine serum (FBS). The second and third aliquots were supplemented with 61.44 µg/mL of the inhibitory protein and 140 or 410 µg/mL of paromomycin, respectively. The fourth and fifth aliquots contained 61.44 µg/mL of the inhibitory protein and 170 or 470 µg/mL of streptomycin, respectively.
Cytotoxicity assay
The cytotoxic effect of the inhibitory protein on the HSF cell line was evaluated using the Sulforhodamine B (SRB) assay.36,37 Approximately 5 × 103 cells in 100 µL of culture medium were seeded into each well of a 96-well plate and incubated for 24 hrs to allow cell attachment. Subsequently, 100 µL of medium containing various concentrations of the inhibitory protein (0.02, 0.2, 2.0, 20, and 200 µg/mL) was added to the wells, with each concentration tested in duplicate. After 72 hrs of exposure, cells were fixed by replacing the medium with 150 µL of 10% trichloroacetic acid (TCA) and incubating at 4 °C for 1 hrs. The plates were then washed five times with distilled water to remove excess TCA. Next, 70 µL of SRB solution (0.4% w/v) was added to each well, and plates were incubated for 10 min in the dark at room temperature. Unbound dye was removed by washing the plates three times with 1% acetic acid, followed by air drying overnight. Bound dye was solubilized with 150 µL of 10 mM Tris buffer, and absorbance was measured at 540 nm using a FLUO star Omega microplate reader (BMG LABTECH, Ortenburg, Germany). Changes in cell morphology after treatment with the inhibitory protein were also examined, including partial or complete loss of the monolayer, cell rounding, shrinkage, and granulation.
Statistical analysis
All experiments were performed in triplicate, and the results are expressed as mean ± standard deviation (SD) and standard error (SE). Data were processed using Microsoft Excel 365 and analyzed with SPSS software version 25. Quantitative data with a normal distribution among different treatments were evaluated using one-way analysis of variance (ANOVA), followed by Tukey’s post hoc test. Differences were considered statistically significant at P < 0.05.
Antibiotic resistance profile of bacterial isolates
Samples of pathogenic bacteria were collected from King Khalid and Al-Salam Hospitals in Riyadh, Saudi Arabia, during January and February 2025. Among the isolates, strain B25 exhibited resistance to amikacin, tobramycin, gentamicin, streptomycin, neomycin, kanamycin, and paromomycin, with minimum inhibitory concentrations (MICs) of 500, 440, 320, 170, 110, 200, and 140 µg/mL, respectively. Strain B75 was resistant to the same antibiotics, with MICs of 80, 140, 200, 470, 440, 500, and 410 µg/mL, respectively. Accordingly, isolate B25 demonstrated the highest resistance to amikacin, tobramycin, and gentamicin, whereas isolate B75 showed the highest resistance to streptomycin, neomycin, kanamycin, and paromomycin (Figure 1). Aminoglycosides remain first-line agents against many Gram-negative and some Gram-positive infections; however, Gram-positive bacteria generally exhibit greater resistance to these antibiotics. Resistance to aminoglycosides occurs through several mechanisms, including restricted uptake through the cell wall, active efflux, modification of target binding sites, and enzymatic inactivation by aminoglycoside-modifying enzymes (AMEs) or related inhibitory proteins. This diversity of resistance mechanisms explains the observed differences in resistance patterns between isolates B25 and B75. The relatively lower MIC values of aminoglycosides compared with the markedly higher MICs of β-lactam antibiotics (penicillins and cephalosporins) confirm that aminoglycosides remain potent agents against many bacterial pathogens, particularly when combined with enzyme inhibitors targeting resistant strains. The obtained MIC values were consistent with previously reported clinical ranges, suggesting that antimicrobial resistance has slightly declined over the past five years. Amikacin inhibited isolates B25 and B75 at 500 and 80 µg/mL, respectively; tobramycin at 440 and 140 µg/mL; and gentamicin at 320 and 200 µg/mL. Hence, isolate B25 was more resistant to these antibiotics than isolate B75, possibly due to synergistic resistance mechanisms unique to B25. Conversely, streptomycin, neomycin, kanamycin, and paromomycin inhibited isolate B75 at 470, 440, 500, and 410 µg/mL, respectively, and isolate B25 at 170, 110, 200, and 140 µg/mL, respectively indicating higher resistance of B75 to these aminoglycosides, likely due to strain-specific properties or enzyme variations. Another 21 bacterial isolates showed mild and inconsistent resistance patterns and were therefore excluded from further analysis.
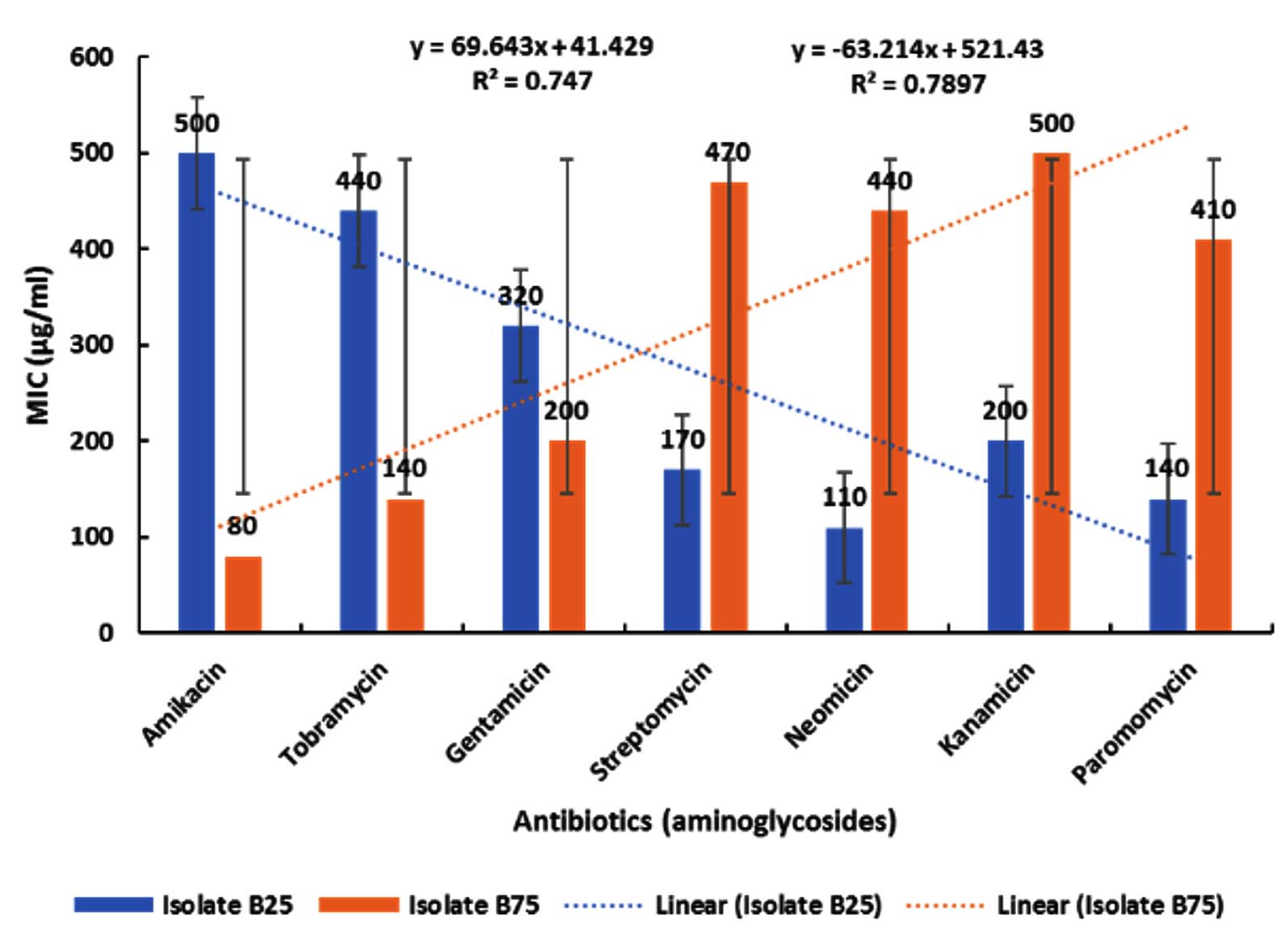
Figure 1. Effect of aminoglycosides on isolates B25 and B75 across increasing concentrations. Data are presented as mean ± standard deviation (SD) of three independent experiments performed in triplicate (n = 9). Statistical analysis was performed using one-way ANOVA followed by Tukey’s post hoc test. A P-value < 0.05 was considered statistically significant. Dotted lines represent linear regression trends
As shown in Figure 1, treatment resulted in a progressive and statistically significant alteration in both measured parameters across the tested concentrations (p < 0.05, one-way ANOVA followed by Tukey’s post hoc test, n = 3 independent experiments performed in triplicate). The blue bars represent mean ± SD of isolate B25, while the orange bars represent mean ± SD of isolate B75. Although variability appears high at certain concentrations, particularly in the mid-treatment groups, statistical analysis confirmed significant differences between treated and control groups (p < 0.05). The increased dispersion observed in some groups reflects biological variability among independent experimental replicates rather than technical inconsistency. Importantly, the overall trend analysis (linear regression) demonstrated a significant negative correlation for isolate B5 (R² = 0.7897, P < 0.05) and a significant positive correlation for isolate B75 (R² = 0.747, P < 0.05), supporting a dose-dependent response despite variability.
Inhibition zone screening assay
Inhibition zone screening revealed that among 88 bacterial isolates, 16 isolates exhibited inhibitory activity against the growth of S. pneumoniae B25 in the presence of amikacin, tobramycin, and gentamicin, and against S. aureus B75 in the presence of streptomycin, neomycin, kanamycin, and paromomycin. The most potent isolate, designated B80, demonstrated the strongest inhibitory effect against all tested aminoglycosides (Table 1). Isolate B80 produced inhibition zones of 42.3, 44.7, 42.3, 40.0, 45.3, 40.0, and 44.7 mm with amikacin, tobramycin, gentamicin, streptomycin, neomycin, kanamycin, and paromomycin, respectively. Streptomycin and kanamycin (40 mm) showed the lowest inhibition, whereas neomycin (45.3 mm) exhibited the highest inhibition, followed by tobramycin and paromomycin (44.7 mm) and amikacin and gentamicin (42.3 mm). Other isolates exhibited moderate inhibitory activity. The B33 isolate showed inhibition zones of 37.6, 36.6, 35.6, 37.3, 36.6, 37.3, and 37.6 mm against amikacin, tobramycin, gentamicin, streptomycin, neomycin, kanamycin, and paromomycin, respectively. Similarly, the B49 isolate exhibited inhibition zones of 30.02, 32.3, 33.3, 35.02, 33.03, 30.02, and 32.03 mm against the same antibiotics. The B56 and B84 isolates showed comparable inhibition patterns, with zones measuring 37.6, 36.6, 35.6, 37.3, 36.6, 37.3, and 37.6 mm against amikacin, tobramycin, gentamicin, streptomycin, neomycin, kanamycin, and paromomycin, respectively. These isolates exhibited identical inhibition against amikacin and paromomycin (37.6 mm), tobramycin and neomycin (36.6 mm), and streptomycin and kanamycin (37.3 mm), while gentamicin showed a distinct inhibition (35.6 mm). The comparable activity against amikacin and kanamycin may be attributed to their structural simila nitrate reductase, urease, Voges-Proskauer rity, as amikacin is derived from kanamycin A by acylation with an L-(-)-γ-amino-α-hydroxybutyryl side chain at the C-1 amino group of the deoxystreptamine nucleus. Streptomycin and kanamycin also share a 2-deoxystreptamine core, which explains their similar inhibitory responses. Isolates B4, B55, and B69 exhibited weaker inhibitory effects, with inhibition zones of 19.0, 22.3, 21.7, 19.3, 19.0, 21.7, and 22.3 mm for amikacin, tobramycin, gentamicin, streptomycin, neomycin, kanamycin, and paromomycin, respectively.
Table (1): Screening of inhibitor-producing bacterial isolates showing inhibitory activity against S. pneumoniae (B25) and S. aureus (B75) in the presence of various aminoglycosides
| Isolates | Mean of inhibition zone diameter (mm) against antibiotics | ||||||
|---|---|---|---|---|---|---|---|
| Am | Tb | Ge | St | Nm | Km | Pm | |
| B4 | 19.03.6SD2.08SE | 22.32.51SD1.45SE | 21.73.51SD2.02SE | 19.34.04SD2.33SE | 19.03.6SD2.08SE | 21.73.51SD2.02SE | 22.32.51SD1.45SE |
| B20 | 30.02.0SD1.15SE | 32.32.51SD1.45SE | 33.33.51SD2.02SE | 35.02.0SD1.15SE | 33.33.51SD2.02SE | 30.02.0SD1.15SE | 32.32.51SD1.45SE |
| B30 | 19.34.04SD2.33SE | 22.32.51SD1.45SE | 19.03.6SD2.08SE | 21.73.51SD2.02SE | 22.32.51SD1.45SE | 19.34.04SD2.33SE | 19.03.6SD2.08SE |
| B33 | 37.62.51SD1.45SE | 36.63.51SD2.02SE | 35.66.02SD3.48SE | 37.32.51SD1.45SE | 36.63.51SD2.02SE | 37.32.51SD1.45SE | 37.62.51SD1.45SE |
| B41 | 19.34.04SD2.33SE | 22.32.51SD1.45SE | 19.03.6SD2.08SE | 21.73.51SD2.02SE | 22.32.51SD1.45SE | 19.34.04SD2.33SE | 19.03.6SD2.08SE |
| B44 | 30.02.0SD1.15SE | 32.32.51SD1.45SE | 33.33.51SD2.02SE | 35.02.0SD1.15SE | 33.33.51SD2.02SE | 30.02.0SD1.15SE | 32.32.51SD1.45SE |
| B49 | 37.62.51SD1.45SE | 36.63.51SD2.02SE | 35.66.02SD3.48SE | 37.32.51SD1.45SE | 36.63.51SD2.02SE | 37.32.51SD1.45SE | 37.62.51SD1.45SE |
| B55 | 19.03.6SD2.08SE | 22.32.51SD1.45SE | 21.73.51SD2.02SE | 19.34.04SD2.33SE | 19.03.6SD2.08SE | 21.73.51SD2.02SE | 22.32.51SD1.45SE |
| B56 | 37.62.51SD1.45SE | 36.63.51SD2.02SE | 35.66.02SD3.48SE | 37.32.51SD1.45SE | 36.63.51SD2.02SE | 37.32.51SD1.45SE | 37.62.51SD1.45SE |
| B62 | 30.02.0SD1.15SE | 32.32.51SD1.45SE | 33.33.51SD2.02SE | 35.02.0SD1.15SE | 33.33.51SD2.02SE | 30.02.0SD1.15SE | 32.32.51SD1.45SE |
| B69 | 19.03.6SD2.08SE | 22.32.51SD1.45SE | 21.73.51SD2.02SE | 19.34.04SD2.33SE | 19.03.6SD2.08SE | 21.73.51SD2.02SE | 22.32.51SD1.45SE |
| B72 | 19.34.04SD2.33SE | 22.32.51SD1.45SE | 19.03.6SD2.08SE | 21.73.51SD2.02SE | 22.32.51SD1.45SE | 19.34.04SD2.33SE | 19.03.6SD2.08SE |
| B80 | 42.32.52SD1.45SE | 44.72.52SD1.45SE | 42.32.52SD1.45SE | 40.02.0SD1.15SE | 45.32.51SD1.45SE | 40.02.0SD1.15SE | 44.72.52SD1.45SE |
| B84 | 37.62.51SD1.45SE | 36.63.51SD2.02SE | 35.66.02SD3.48SE | 37.32.51SD1.45SE | 36.63.51SD2.02SE | 37.32.51SD1.45SE | 37.62.51SD1.45SE |
| B85 | 19.34.04SD2.33SE | 22.32.51SD1.45SE | 19.03.6SD2.08SE | 21.73.51SD2.02SE | 22.32.51SD1.45SE | 19.34.04SD2.33SE | 19.03.6SD2.08SE |
| B88 | 30.02.0SD1.15SE | 32.32.51SD1.45SE | 33.33.51SD2.02SE | 35.02.0SD1.15SE | 33.33.51SD2.02SE | 30.02.0SD1.15SE | 32.32.51SD1.45SE |
Am: Amikacin, Tb: Tobramycin, Ge: Gentamicin, St: Streptomycin, Nm: Neomycin, Km: Kanamycin, Pm: Paromomycin, SD: Standard deviation, SE: Standard error
These isolates showed identical inhibition on amikacin and neomycin (19 mm), tobramycin and paromomycin (22.3 mm), and gentamicin and kanamycin (21.7 mm), with streptomycin displaying a distinct effect (19.3 mm). Although amikacin is a semi-synthetic derivative of kanamycin, its lower inhibition zone suggests higher efficacy of the natural antibiotic. Similarly, B20, B44, B62, and B88 produced inhibition zones of 30.0, 32.3, 33.3, 35.0, 33.3, 30.0, and 32.3 mm for the respective antibiotics, showing identical inhibition for amikacin and kanamycin (30 mm), tobramycin and paromomycin (32.3 mm), gentamicin and neomycin (33.3 mm), and a distinct effect for streptomycin (35 mm). Another group, B30, B41, B72, and B85 displayed the lowest inhibitory activity, with inhibition zones of 19.3, 22.3, 19.0, 21.7, 22.3, 19.3, and 19.0 mm, respectively. These isolates showed similar inhibition for amikacin and kanamycin (19.3 mm), tobramycin and neomycin (22.3 mm), and gentamicin and paromomycin (19.0 mm), with a specific inhibition for streptomycin (21.7 mm). Aminoglycosides constitute a broad class of antibiotics containing an aminocyclitol nucleus (streptamine, 2-deoxystreptamine, or streptidine) linked to amino sugars by glycosidic bonds, or in some cases, lacking these sugars as in spectinomycin.
Although mainly used to treat Gram-negative infections, aminoglycosides are also effective against Gram-positive bacteria when combined with β-lactams, vancomycin, or modifying enzyme inhibitors (inhibitory proteins). Streptomycin, the first discovered aminoglycoside, remains clinically significant for Gram-positive infections such as tuberculosis when used in combination therapy. Amikacin and kanamycin, second-line aminoglycosides, are active against a broad spectrum of pathogens, including Mycobacterium tuberculosis, Yersinia pestis, Francisella tularensis, Brucella spp., and P. aeruginosa. Actinomycetes are the primary natural producers of aminoglycosides, which have since evolved into semi-synthetic derivatives with improved pharmacological properties. The most clinically important members include neomycin, kanamycin, gentamicin, netilmicin, tobramycin, and amikacin, produced by Streptomyces fradiae, S. kanamyceticus, Micromonospora purpurea, S. sisomicin, S. tenebrarius, and S. kanamyceticus, respectively. Aminoglycosides demonstrate potent activity against Gram-negative bacteria such as E. coli, Klebsiella pneumoniae, Enterobacter cloacae, Proteus spp., Serratia spp., P. aeruginosa, and Acinetobacter baumannii, while synergistic combinations with other antimicrobials enhance efficacy against Gram-positive pathogens including MRSA, VRSA, VISA, and Mycobacterium species. AMEs, typically encoded on plasmids, are a key resistance mechanism in both Gram-positive and Gram-negative bacteria. Over 100 AMEs have been identified and are classified into three groups: N-acetyltransferases (AACs), O-phosphotransferases (APHs), and O-nucleotidyltransferases (ANTs) based on their ability to acetylate, phosphorylate, or adenylate amino or hydroxyl groups on the aminoglycoside scaffold. These enzymatic modifications reduce antibiotic binding affinity to ribosomal targets, leading to a loss of antibacterial activity.
Identification of bacterial isolates
Isolate B25, which exhibited the highest resistance to amikacin, tobramycin, and gentamicin, and isolate B75, which showed the greatest resistance to streptomycin, neomycin, kanamycin, and paromomycin, were identified as diploid Gram-positive cocci. Microscopic examination revealed that isolate B25 displayed the typical morphology of Streptococcus spp. (Figure 2A), whereas isolate B75 exhibited the characteristic appearance of Staphylococcus spp. (Figure 2B). Biochemical characterization showed that isolate B25 tested negative for catalase, oxidase, urease, alkaline phosphatase, motility, and Voges–Proskauer tests, and did not ferment mannitol, ribose, or sorbitol. However, it tested positive for bile solubility and β-hemolysis, and was able to ferment arabinose, fructose, galactose, glucose, glycerol, glycogen, inulin, lactose, maltose, raffinose, starch, sucrose, and xylose. In contrast, isolate B75 tested negative for gas and hydrogen sulfide production, indole, oxidase, and motility, and did not ferment raffinose, ribose, or xylose. It tested positive for catalase, coagulase, citrate, gelatinase, nitrate reductase, urease, Voges-Proskauer, α-hemolysis, alkaline phosphatase, arginine dehydrolase, and lipase, and was capable of fermenting glucose, galactose, fructose, lactose, maltose, mannitol, mannose, starch, and sucrose. Based on these morphological and biochemical results, which correspond with the descriptions in Bergey’s Manual of Determinative Bacteriology (1989), isolate B25 was identified as S. pneumoniae, and isolate B75 as S. aureus.
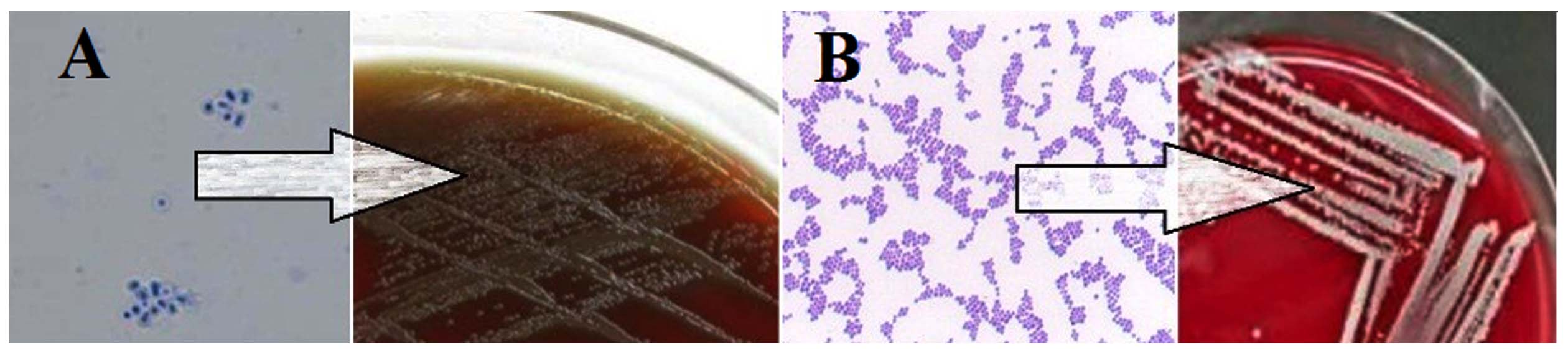
Figure 2. Microscopic and cultural characteristics of the bacterial isolates. (A) Gram-stained smear observed under 100× oil immersion objective (1000× total magnification). Scale bar = 10 µm. (B) Gram-stained smear under identical magnification. Scale bar = 10 µm
Partial DNA sequencing of isolate B25 was performed using universal primers for the 16S rRNA gene. The adjacent fragment (1500 bp) was generated from the isolate using Codon-Code Aligner software. Next, the closest reference sequences for the species and subspecies were imported from GenBank. The phylogenetic tree was constructed using the Neighbor-Joining method with 1000 bootstrap values. Isolate B25 was found to belong to the Streptococcaceae family and was closely related to members of the genus Streptococcus (Figure 3A), and was found to belong to 3 clusters: cluster 1 contains S. pneumoniae (LC720404.1), cluster 2 contains S. pneumoniae (KF815703.1), (EU170480.1), and (KR349544), and cluster 3 contains S. pneumoniae (MW301141.1) and (EU834943.1), with 99% sequence similarity. The same technique was used with isolate B75, which had a 1550 bp contiguous fragment generated using the Codon-Code Aligner software. Isolate B75 was found to belong to the Staphylococcaceae family and is closely related to members of the genus Streptococcus (Figure 3B), and was found to belong to 3 clusters: cluster 1 contains S. aureus (PQ141662), cluster 2 contains S. aureus (PQ141655), (PQ141656), and (PQ141657), and cluster 3 contains S. aureus (PQ141660) and (PQ141659), with 99% sequence similarity. Partial DNA sequencing of isolate B25 was conducted using universal primers targeting the 16S rRNA gene. A contiguous fragment of approximately 1500 bp was assembled using CodonCode Aligner software. The closest reference sequences at the species and subspecies levels were retrieved from GenBank, and a phylogenetic tree was constructed using the Neighbor-Joining method with 1000 bootstrap replications. Isolate B25 was identified as a member of the Streptococcaceae family, showing close affiliation with species of the genus Streptococcus (Figure 3A). It clustered into three groups: Cluster 1 included S. pneumoniae (LC720404.1); Cluster 2 included S. pneumoniae (KF815703.1, EU170480.1, KR349544.1); and Cluster 3 included S. pneumoniae (MW301141.1, EU834943.1), all sharing 99% sequence similarity. Similarly, partial sequencing of isolate B75 produced a contiguous fragment of approximately 1550 bp using CodonCode Aligner. Phylogenetic analysis placed isolate B75 within the Staphylococcaceae family, closely related to species of the genus Staphylococcus (Figure 3B). The isolate formed three clusters: Cluster 1 included S. aureus (PQ141662); Cluster 2 included S. aureus (PQ141655, PQ141656, PQ141657); and Cluster 3 included S. aureus (PQ141660, PQ141659), with 99% sequence similarity.

Figure 3. Phylogenetic tree of genetic identification: (A) B25 S. pneumoniae as the most resistant isolate to amikacin, tobramycin, and gentamicin, (B) B75 S. aureus as the most resistant isolate to streptomycin, neomycin, kanamycin, and paromomycin
Isolate B80, identified as the most prolific producer of the modifying enzyme inhibitor, was observed to be a Gram-negative bacillus (Figure 4A). Based on biochemical characterization, isolate B80 exhibited negative results for coagulase, gas and hydrogen sulfide production, indole, methyl red, urease, Voges-Proskauer, esculin hydrolysis, and lecithinase tests, and was unable to ferment galactose, glucose, lactose, inulin, maltose, mannose, sorbitol, starch, sucrose, and xylose. Conversely, the isolate showed positive reactions for catalase, citrate, cetrimide, gelatinase, β-hemolysis, motility, nitrate reductase, oxidase, acetate utilization, and lipase, and was able to ferment fructose, malonate, mannitol, and ribose.
Partial 16S rRNA gene sequencing of isolate B80 was carried out using universal primers, yielding an approximately 1500 bp fragment assembled with CodonCode Aligner software. The most closely related reference sequences were retrieved from GenBank, and a phylogenetic tree was constructed using the Neighbor-Joining method with 1000 bootstrap replications. Isolate B80 was identified as a member of the Pseudomonadaceae family, showing a close phylogenetic relationship to species of the genus Pseudomonas (Figure 4B). The isolate clustered into three groups: Cluster 1 included P. aeruginosa (JQ041638); Cluster 2 included P. aeruginosa (JQ659978, JF708942, JQ659819); and Cluster 3 included P. aeruginosa (JQ659717, JQ659921), all sharing 99% sequence similarity.
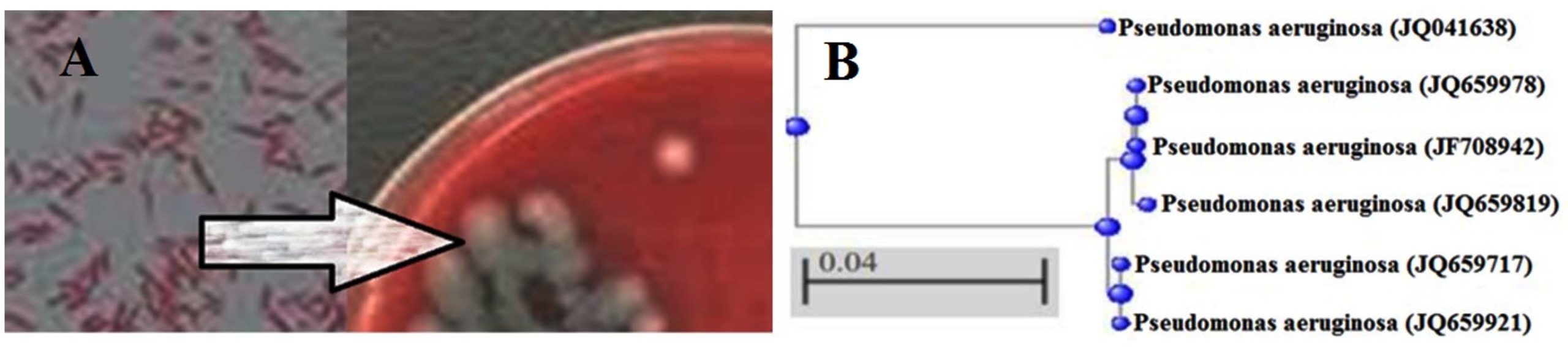
Figure 4. Identification of the bacterial isolate. (A) Gram-stained smear showing Gram-negative rod-shaped cells observed under oil immersion (100× objective; 1000× total magnification). Scale bar = 10 µm. (B) Phylogenetic tree based on 16S rRNA gene sequences showing the relationship between the studied isolate and reference P. aeruginosa strains. The tree was constructed using the neighbor-joining method. The scale bar (0.04) represents nucleotide substitutions per site
Optimization of environmental conditions
P. aeruginosa (B80) exhibited the highest production of a modifying enzyme inhibitor active against both S. pneumoniae (B25) (in the presence of amikacin, tobramycin, and gentamicin) and S. aureus (B75) (in the presence of streptomycin, neomycin, kanamycin, and paromomycin). The activity of the modifying enzyme inhibitor was optimized under various environmental conditions, including incubation period, temperature, pH, stirring speed, aeration rate, and different carbon and nitrogen sources, using submerged fermentation. Maximum inhibitor activity was achieved after 36 hrs (Figure 5A), at 35 °C (Figure 5B), pH 7.4 (Figure 5C), 140 rpm (Figure 5D), and 60% aeration (Figure 5E), with glucose and ammonium sulfate serving as the optimal carbon and nitrogen sources, respectively (Figures 5F and 5G). P. aeruginosa (B80), identified as a mesophilic and metabolically versatile pathogen, demonstrated optimal growth and inhibitor production at moderate temperatures (25-40 °C), with the peak at 35 °C after 36 h of incubation, coinciding with maximal production of primary and secondary metabolites. A neutral pH (7.4) favored both bacterial growth and enzyme inhibitor production, while acidic or alkaline conditions reduced activity. Although acidic environments can induce adaptive physiological and genetic responses in P. aeruginosa that enhance virulence and antibiotic resistance, they markedly decreased the activity of the modifying enzyme inhibitor. Stirring speed and aeration significantly influenced enzyme inhibitor production. Low agitation reduced oxygen availability and transport, thereby limiting growth and metabolism, whereas excessive agitation caused cellular stress and mechanical damage, leading to reduced viability. The optimal stirring speed (140 rpm) supported balanced oxygen transfer and metabolic activity. Similarly, moderate aeration (60%) was found to be optimal; while P. aeruginosa requires oxygen as a terminal electron acceptor, excessive aeration caused oxidative stress and cell death.
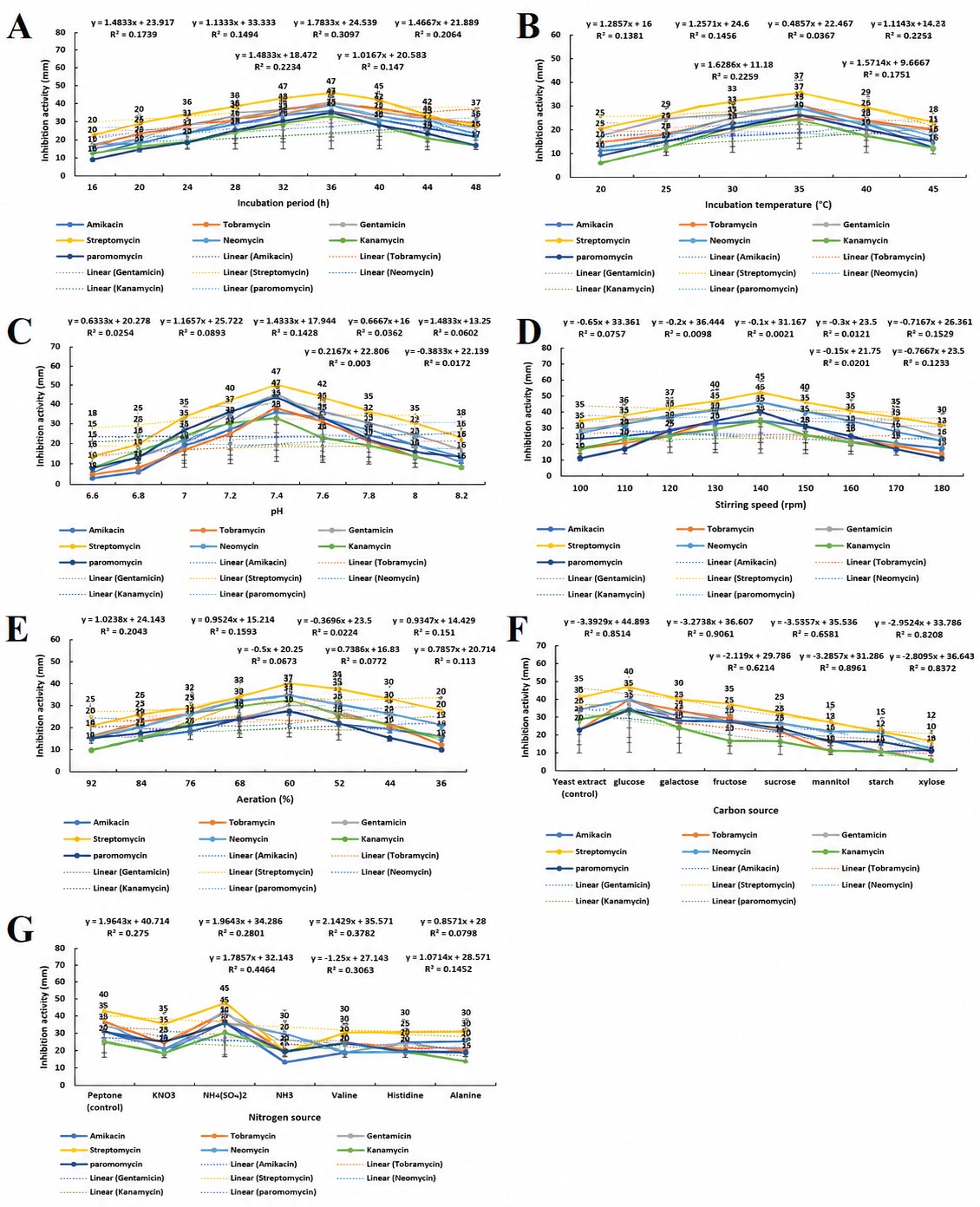
Figure 5. Optimization of modifying enzyme inhibitor activity produced by P. aeruginosa (B80) under varying environmental conditions. Effects of (A) incubation period, (B) incubation temperature, (C) pH, (D) stirring speed, (E) aeration, (F) carbon source, and (G) nitrogen source on the inhibition of S. pneumoniae (B25) and S. aureus (B75) in the presence of aminoglycosides are shown. Values represent the mean ± SD of triplicate experiments
Among carbon sources tested, glucose yielded the highest inhibitor activity, outperforming yeast extract and other sugars. Glucose provides an easily metabolizable energy source that supports cell growth, biofilm formation, and synthesis of key extracellular metabolites. Ammonium sulfate proved to be the most effective nitrogen source, surpassing peptone and other nitrogenous compounds. Its use enhanced cyanide production via RhlR-regulated pathways, maintaining cooperative bacterial populations and metabolic balance. Streptomycin showed the highest activity with the modifying enzyme inhibitor under optimal conditions, with inhibition zones of 50, 37, 47, 45, 37, 42, and 45 mm at optimal incubation time, temperature, pH, stirring speed, aeration, carbon source, and nitrogen source, respectively. At the optimal incubation period, gentamicin, tobramycin, neomycin, amikacin, paromomycin, and kanamycin showed activities of 45, 44, 43, 40, 39, and 37 mm, respectively. At the optimal temperature, the corresponding activities were 32, 30, 27, and 25 mm; at the optimal pH, 42, 40, 37, 35, and 32 mm; at the optimal stirring speed, 40, 35, and 30 mm; at the optimal aeration, 37, 32, 30, and 28 mm; at the optimal carbon source, 35 and 30 mm; and at the optimal nitrogen source, 40, 35, and 30 mm. These results demonstrate that the optimized fermentation conditions significantly enhanced inhibitor activity, with streptomycin exhibiting the highest synergistic effect among the tested aminoglycosides.
Purification and electrophoresis of modifying enzyme inhibitor
A modifying enzyme inhibitor with peak activity was produced by isolate B80 under optimized conditions and purified through a stepwise process, including precipitation with saturated ammonium sulfate, dialysis, ion-exchange chromatography, gel filtration chromatography, and final dialysis to concentrate the protein. The inhibitor was precipitated using 70% saturated ammonium sulfate, resulting in activity of 192 U/mL, specific activity of 1.4 U/mg, a purification fold of 2.4, and a yield of 28.5% relative to the crude lysate (675 U/mL, 0.58 U/mg, 1.0-fold, 100% yield). This step increased the specific activity from 0.58-1.4 U/mg, indicating the removal of harmful impurities. Further purification via ion-exchange and gel filtration chromatography increased the specific activity to 3.0 and 4.5 U/mg, corresponding to 7.8-fold purification and 46.7% yield (Table 2). SDS-PAGE analysis revealed a single protein band at approximately 20 kDa, confirming the purity of the modifying enzyme inhibitor (Figure 6). The inhibitor’s small size and ability to precipitate at high ammonium sulfate concentrations is consistent with the “salting out” effect, where salt ions compete with protein molecules for water, reducing protein solubility and promoting aggregation and precipitation.
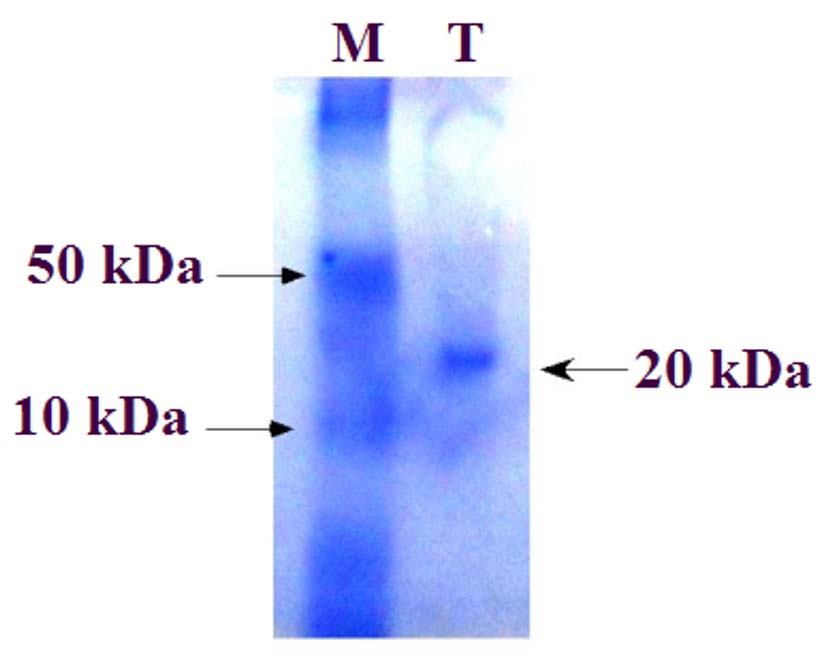
Figure 6. SDS-PAGE analysis of the purified modifying enzyme inhibitor produced by P. aeruginosa (B80). A single protein band was observed at approximately 20 kDa, confirming the purity of the inhibitor
Table (2): Purification table of modifying enzyme inhibitor
(NH4)2SO4 concen. |
Total activity (U) |
Total protein (mg) |
Specific activity (U/mg) |
Purification fold |
Yield (%) |
|---|---|---|---|---|---|
Lysate |
675 |
1174 |
0.58 |
1.00 |
100 |
30% |
0.00 |
85 |
0.00 |
0.00 |
0.00 |
40% |
0.00 |
92 |
0.00 |
0.00 |
0.00 |
50% |
0.00 |
105 |
0.00 |
0.00 |
0.00 |
60% |
0.00 |
120 |
0.00 |
0.00 |
0.00 |
70% |
192 |
135 |
1.4 |
2.4 |
28.5 |
80% |
0.00 |
115 |
0.00 |
0.00 |
0.00 |
90% |
0.00 |
95 |
0.00 |
0.00 |
0.00 |
DEAE-Cellulose |
247 |
82 |
3.00 |
5.2 |
36.6 |
Sephadex G100 |
315 |
70 |
4.5 |
7.8 |
46.7 |
Synergistic effect of antibiotics and modifying enzyme inhibitor
Aminoglycoside antibiotics exhibited variable MICs against S. pneumoniae (B25) and S. aureus (B75). Therefore, a wide range of modifying enzyme inhibitor concentrations (0.015-245.76 µg/mL) was tested in combination with the MICs of each antibiotic (Figure 7A, 7B). Against S. pneumoniae (B25), 61.44 µg/mL of modifying enzyme inhibitor was most effective with all antibiotic MICs. At this concentration, paromomycin (140 µg/mL) produced the highest inhibition (57 mm), followed by streptomycin (170 µg/mL, 52 mm), amikacin (500 µg/mL) and gentamicin (320 µg/mL, 50 mm), kanamycin (200 µg/mL, 48 mm), neomycin (110 µg/mL, 44 mm), and tobramycin (440 µg/mL, 40 mm). Gentamicin was slightly more effective than amikacin when combined with the inhibitor. The inhibitor was active at very low concentrations (from 0.015 µg/mL), peaked at 61.44 µg/mL, and declined at higher concentrations, likely due to self-binding, substrate-level inhibition, or structural interference with the target enzyme. Similarly, against S. aureus (B75), 61.44 µg/mL of modifying enzyme inhibitor was optimal. Paromomycin (410 µg/mL) showed the highest inhibitory activity (60 mm), followed by streptomycin (470 µg/mL, 57 mm), kanamycin (500 µg/mL, 55 mm), neomycin (440 µg/mL, 48 mm), tobramycin (140 µg/mL, 42 mm), gentamicin (200 µg/mL, 38 mm), and amikacin (80 µg/mL, 35 mm). These results indicate that paromomycin and streptomycin are the most effective aminoglycosides when combined with 61.44 µg/mL of modifying enzyme inhibitor, whereas other aminoglycosides showed variable activity. This study recommends the use of paromomycin and streptomycin with the modifying enzyme inhibitor at the indicated concentrations to effectively inhibit resistant S. pneumoniae (B25) and S. aureus (B75), which are major causes of severe Gram-positive infections.
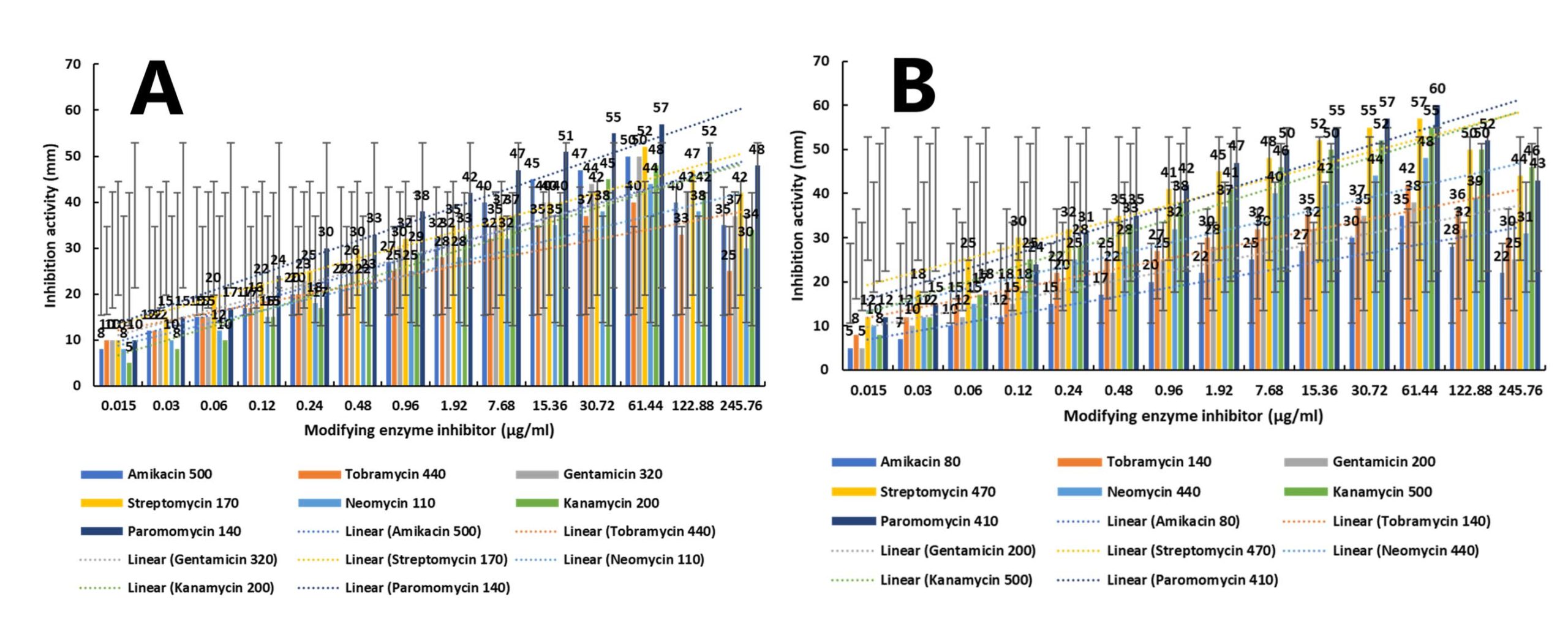
Figure 7. Dose-dependent enhancement of antibiotic inhibition activity following treatment with modifying enzyme inhibitor. (A) High initial antibiotic concentration panel. (B) Lower initial antibiotic concentration panel. Data represent mean ± SD (n = 3). Dotted lines indicate linear regression trends
Cytotoxicity of modifying enzyme inhibitor on human skin fibroblast (HSF) cell line
The cytotoxicity of the modifying enzyme inhibitor against HSF cells was evaluated using the SRB assay. Cells were treated with paromomycin (140 and 410 µg/mL), streptomycin (170 and 470 µg/mL), and the inhibitor (61.44 µg/mL) for 24 hrs. HSF cell viability remained approximately 99%, with cytotoxicity below 5%, and no morphological changes, including rounding, shrinkage, or granulation, were observed compared to the negative control (Figure 8A). These results indicate that formulations of paromomycin (Figure 8B, 8C) or streptomycin (Figure 8D, 8E) combined with the modifying enzyme inhibitor are safe for human skin and do not cause significant adverse effects. The low toxicity of the inhibitor supports high cell availability by minimizing off-target interactions and facilitating skin penetration and retention. Moreover, chemical modifications of the inhibitor enhance specificity for the target enzyme, further reducing potential cellular toxicity. Cytotoxicity assays are essential for identifying safe, biocompatible compounds and evaluating the suitability of medical materials. In vitro testing is preferred over in vivo studies due to reproducibility, reduced animal usage, and easier access to materials. The IC50, representing the concentration that reduces cell viability by 50%, is commonly used to quantify cytotoxic effects.
![]()
Figure 8. Morphological changes observed under light/phase-contrast microscopy. (A) Untreated control showing intact cellular morphology. (B-E) Cells exposed to increasing concentrations of aminoglycoside and modifying-enzyme inhibitor, demonstrating progressive structural alterations. Images were captured at 400× total magnification. Scale bar = 10 µm
Although slight variations in cell density appearance were observed in Figure 8B and 8E, quantitative viability analysis demonstrated that all treatments maintained high cell survival rates (98.7%-99.3%), with no statistically significant difference compared to the untreated control (one-way ANOVA, P > 0.05). Colony-forming unit (CFU) counts showed comparable values between treated and control groups (log CFU/mL difference < 0.05). Furthermore, no structural abnormalities such as membrane rupture, cytoplasmic leakage, or cell shrinkage were detected in the high-resolution images. The visual perception of reduced density in some panels was attributed to minor differences in focal plane and field selection rather than actual cytotoxic effects.
Multidrug-resistant pathogenic bacteria exhibit variable resistance rates against aminoglycosides, including amikacin, tobramycin, gentamicin, streptomycin, neomycin, kanamycin, and paromomycin, reflected by differing MICs. Although all these antibiotics belong to the aminoglycoside class, each displays a specific MIC against various bacterial isolates, particularly the highly resistant S. pneumoniae (B25) and S. aureus (B75). Despite the notable nephrotoxicity of broad-spectrum aminoglycosides, they are widely used in combination with modifying enzyme inhibitors to combat resistant pathogens. Ribostamycin, a potent aminoglycoside, exhibits MICs ranging from 0.9-7.2 µM against three E. coli strains and 0.5 µM against Haemophilus influenzae. When combined with ethylenediaminetetraacetic acid (EDTA), these MICs improve up to 8-fold, ranging from 57.2-7.2 µM. Aminoglycosides such as gentamicin, amikacin, netilmicin, isepamicin, and ribostamycin have been tested against Gram-negative bacteria (E. coli ATCC 25922, P. aeruginosa ATCC 27853, H. influenzae ATCC 49247) and Gram-positive bacteria (S. epidermidis ATCC 12228, S. aureus ATCC 12600, E. faecalis ATCC 29212, S. pneumoniae ATCC 49619). All aminoglycosides were highly active against H. influenzae, S. epidermidis, and S. pneumoniae, with MIC values ranging from 0.1-13.7 µM. Netilmicin demonstrated broad efficacy, with MICs ranging from 0.1-5.6 µM. Ribostamycin exhibited moderate activity against E. coli (MIC 29 µM), very weak activity against P. aeruginosa, S. aureus, and E. faecalis (MIC <115.8 µM), and very potent activity against H. influenzae and S. pneumoniae (MIC 0.5 and 7.2 µM, respectively).38 Although E. faecalis is a common intestinal commensal, it has been implicated in recurrent urinary tract infections and bacteremia.39 S. aureus, a member of the normal human microflora, acts as an opportunistic pathogen causing infections of the skin, soft tissues, respiratory system, and urinary tract.40 MIC values of aminoglycosides against these pathogens were determined using an inoculum of 5 × 105 CFU/mL. Ribostamycin has emerged as a promising aminoglycoside for treating resistant Gram-positive and Gram-negative infections, especially E. coli and Staphylococcus spp. EDTA, by contrast, exhibits a different mechanism: it chelates divalent cations such as magnesium, increasing cell membrane permeability. This enhanced permeability allows aminoglycosides to penetrate bacterial cells more effectively and inhibit protein synthesis. However, EDTA does not inhibit modifying enzymes directly; it solely facilitates antibiotic entry to potentiate their antibacterial activity.41
The relatively large standard deviations observed in some treatment groups likely reflect intrinsic biological variability associated with microbial adaptive responses under stress conditions. Such variability has been widely reported in studies involving antimicrobial exposure and epigenetic modulation, where heterogeneous population responses may occur due to differential gene expression or resistance-associated mechanisms. Importantly, despite the observed dispersion, statistical analysis confirmed significant differences between experimental groups (P < 0.05), indicating that the treatment effect remains robust. The variability was more pronounced at intermediate concentrations, which may represent transitional physiological states where subpopulations exhibit differential sensitivity. Furthermore, the consistent directional trends decreasing trend in isolate B25 and increasing trend in isolate B75 and significant regression coefficients support the biological relevance of the findings. Therefore, the magnitude of error bars does not undermine the statistical or biological significance of the results but rather reflects expected biological heterogeneity.
Out of 88 bacterial isolates, 16 isolates showed inhibition of S. pneumoniae B25 and S. aureus B75 using amikacin, tobramycin, gentamicin, streptomycin, neomycin, kanamycin, and paromomycin. The most efficient isolate for producing a modifying enzyme inhibitor was identified as P. aeruginosa B80. Although S. pneumoniae B25, S. aureus B75, and P. aeruginosa B80 are pathogenic bacteria, antagonism has been demonstrated between the latter isolate and the former two, with P. aeruginosa B80 producing an inhibitor that inhibits a modifying enzyme used by S. pneumoniae B25 and S. aureus B75 to modify and inactivate aminoglycosides, leading to resistance development. Colonic bacteria, including E. coli, K. pneumoniae, and E. aerogenes were inhibited by P. aeruginosa in antagonism, especially with P. aeruginosa producing pyocyanin and/or pyoverdin.42 E. aerogenes and E. coli were inhibited in antagonism by P. aeruginosa, and E. aerogenes was found to be more sensitive. These results show the same pattern, with Ea1 (n = 22) being the most sensitive, followed by Ec2 (n = 21). Ec1 (n = 12) and Kp1 (n = 15) were less sensitive to P. aeruginosa. These results show that coliform isolates from the P. aeruginosa-free sample were more susceptible than isolates from the P. aeruginosa-containing sample, indicating the acquisition of resistance by these bacteria.43 Pseudomonas spp. showed antagonistic effect on foodborne spoilage and pathogenic bacteria. Thus, 13 Pseudomonas strains showed inhibition of other Pseudomonas spp., including P. phaseolicola, P. pisi, P. putida, P. fluorescens, and P. fragi, and showed antagonism to non-pseudomonads, including S. aureus. P. aeruginosa antagonizes S. pneumoniae by producing protease IV (PIV) which damages host lung tissue and increases inflammation caused by S. pneumoniae infection.44 This interaction also involves a disruption of the host’s immune response, creating a favorable environment for P. aeruginosa. While S. pneumoniae do not directly antagonize P. aeruginosa, which can inhibit the growth of S. pneumoniae, and there is evidence that S. pneumoniae inhibit P. aeruginosa in laboratory models.45 P. aeruginosa antagonizes S. aureus by secreting proteases (LasA, LasB), siderophores, and the quinolone HQNO, which inhibit S. aureus metabolism. HQNO inhibits the electron transport chain in S. aureus, converting aerobic respiration to fermentation in metabolism, and reducing cell viability. Siderophores such as pyoverdine and pyochelin also convert aerobic respiration into fermentation in the metabolism of S. aureus. Proteases, including LasA, LasB, and AprA, degrade S. aureus surface proteins such as SasG, leading to cell lysis. Furthermore, P. aeruginosa type IV invades S. aureus colonies, inactivating them and increasing their susceptibility to antibiotics. Furthermore, polysaccharide-producing P. aeruginosa antagonizes S. aureus.46
Amikacin, tobramycin, and gentamicin exhibited high inhibitory activity against most bacterial isolates, except for S. pneumoniae (B25), which demonstrated high resistance. Conversely, S. aureus (B75) showed the greatest resistance to streptomycin, neomycin, kanamycin, and paromomycin. S. aureus infections, particularly MRSA strains, are widespread in hospitals, accounting for approximately 18%-28% of cases. Methicillin-sensitive S. aureus (MSSA) strains can acquire resistance genes through genetic mutation, evolving into MRSA strains. These MRSA strains exhibit increased sensitivity to various antibiotic classes when combined with enzyme inhibitors, including aminoglycosides, which alone have limited effectiveness.47 Mutated MRSA strains also display high resistance to quinolones, contributing to widespread distribution and severe infections. Resistant S. pneumoniae strains are prevalent globally and cause diverse respiratory infections. Multidrug-resistance is common, particularly against ampicillin (6%-17%) and erythromycin (33%) through the MLS (macrolides, lincosamides, streptogramin B) mechanism. Combining enzyme inhibitors with antibiotics is necessary to inhibit these resistant bacteria, which often harbor multiple resistance mechanisms, posing a significant medical challenge. Gram-positive cocci, including S. aureus, Enterococcus spp., and S. pneumoniae, are among the most common resistant pathogens, frequently carrying resistance genes on plasmids.48 Although S. aureus generally shows low resistance to gentamicin, mutant strains particularly in nosocomial infections exhibit high resistance. Aminoglycosides remain widely prescribed for serious infections, especially in combination with enzyme inhibitors, which enhance their efficacy. S. pneumoniae has also evolved multiple resistance mechanisms to circumvent the inhibitory effects of diverse antibiotic classes, including β-lactams, macrolides, lincosamides, sulfonamides, and fluoroquinolones.49
P. aeruginosa B80 isolate was found to have the highest production of modifying enzyme inhibitor against both S. pneumonia B25 with amikacin, tobramycin, and gentamicin, and S. aureus B75 with streptomycin, neomycin, kanamycin, and paromomycin. The activity of modifying enzyme inhibitor was improved in accordance with the optimization of environmental conditions, including incubation period, incubation temperature, pH, stirring speed, aeration, and carbon and nitrogen sources through submerged fermentation. Peak activity was obtained at 36 hrs, 35 °C, pH 7.4, 140 rpm, 60% aeration, and glucose and ammonium sulfate as the best carbon and nitrogen sources, respectively, with amikacin, tobramycin, and gentamicin against S. pneumoniae B25, and with streptomycin, neomycin, kanamycin, and paromomycin against S. aureus B75. The maximum activity of modifying enzyme inhibitor using gentamicin was obtained when the fermentation medium (pH 7.5) containing glucose and asparagine as carbon and nitrogen sources, respectively, was inoculated with P. aeruginosa (8 ×107 CFU/ml) and incubated at 30 °C for 24 hrs.50,51 The activity of aminoglycoside modifying enzymes and their inhibitors is greatly influenced by incubation temperature, with temperatures above 35 °C reducing or inhibiting the activity of most metabolites, including enzymes and enzyme inhibitors. However, thermotolerant bacteria showed maximum metabolic activity at high temperatures consistent with physiological nature. Aminoglycosides are first-line antibiotics for treating many severe infections, but they have lost significant effectiveness over the past 20 years due to the development of bacterial resistance. Aminoglycoside resistance develops through several mechanisms, including enzymatic modification. Aminoglycosides contain a 2-deoxystreptamine core containing hydroxyl and amine groups, which can be modified by modifying enzymes, including nucleotidyl transferases, phosphotransferases, and acetyltransferases. These enzymes can be inhibited by specific proteins, allowing aminoglycosides to block protein synthesis and prevent bacterial growth.52
A modifying enzyme inhibitor was precipitated using 70% saturated ammonium sulfate, yielding a total activity of 192 U/mL, a specific activity of 1.4 U/mg, a purification fold of 2.4, and a recovery yield of 28.5%, compared to the crude lysate values of 675 U/mL, 0.58 U/mg, 1.0, and 100%, respectively. SDS-PAGE analysis of the purified inhibitor revealed a single distinct protein band corresponding to an approximate molecular weight of 20 kDa, confirming the purity of the preparation. Similarly, the successful precipitation of bacteriocin using 70% saturated ammonium sulfate, which enhanced its total activity from 320-5120 U/mL, with a 32% yield and a 3.48-fold purification increase.53 In comparison to ethyl acetate extraction, ammonium sulfate precipitation more effectively retained bacteriocin activity and produced a higher yield.54 Further purification of bacteriocin using cation-exchange chromatography revealed two absorption peaks, of which only the second exhibited antibacterial activity, indicating that the compound was a cationic peptide.55 Concentration of the active fractions increased bacteriocin titers to 5120 U/mL with a 16% yield and a 16.68-fold purification, while subsequent purification by Superdex G-75 gel filtration yielded a single active peak with a total activity of 10,240 U/mL, a 16% yield, and a 34.37-fold purification.
Synergistic testing revealed that paromomycin at 140 and 410 µg/mL against S. pneumoniae (B25) and S. aureus (B75), respectively, exhibited the highest inhibitory activity when combined with 61.44 µg/mL of the modifying enzyme inhibitor, followed by streptomycin at 170 and 470 µg/mL. Similarly, complete inhibition of resistant S. aureus strains using a combination of 128 mg/L of a modifying enzyme inhibitor and 128 mg/L of gentamicin.50 Antibiotic resistance remains a major global health concern, particularly among Gram-positive and Gram-negative pathogens resistant to last-line therapies. Aminoglycosides, which act by binding to the bacterial 30S ribosomal subunit and inducing mRNA misreading, have regained scientific interest due to their potent bactericidal activity and potential for synergism with other agents.56 However, their clinical efficacy is frequently undermined by enzymatic modification, efflux mechanisms, and ribosomal mutations.
To overcome these limitations, recent strategies have focused on combining aminoglycosides with inhibitory proteins or protein-targeting agents that enhance drug uptake, suppress resistance mechanisms, or synergistically disrupt protein synthesis. Aminoglycosides induce protein misfolding and aggregation, leading to cell death through reactive oxygen species (ROS) generation and proteotoxic stress.57 Inhibiting bacterial chaperones or proteases responsible for protein refolding, such as Hsp70 and Clp proteases, has been shown to potentiate aminoglycoside activity by amplifying misfolded protein accumulation.58 Additionally, antimicrobial peptides (AMPs) such as PR-39 and LL-37 enhance aminoglycoside efficacy by permeabilizing bacterial membranes and disrupting intracellular processes, including protein translation.59 Engineered arginine-rich peptide conjugates of aminoglycosides also exhibit improved cellular uptake and dual translation inhibition mechanisms, supporting the development of novel hybrid antimicrobials.60
Another promising strategy targets aminoglycoside-modifying enzymes using kinase inhibitors, as these enzymes share structural similarities with eukaryotic kinases. Compounds such as tyrphostins and staurosporine analogs effectively inhibit aminoglycoside phosphotransferases, restoring antibiotic activity in resistant strains.61 Furthermore, inhibitors of translation elongation factors (e.g., EF-Tu, EF-G) or ribosome recycling proteins can act synergistically with aminoglycosides by simultaneously disrupting multiple stages of protein synthesis. Despite these promising findings, clinical translation of protein-based inhibitors remains challenging due to stability, delivery, and immunogenicity concerns. Advances in nanoparticle-based delivery systems, protein engineering, and conjugate development are likely to overcome these barriers, paving the way for next-generation aminoglycoside-protein inhibitor combinations capable of breaking resistance and restoring antibiotic efficacy.
The four in vitro therapeutic formulations, paromomycin (140 µg/mL + modifying enzyme inhibitor 61.44 µg/mL), paromomycin (410 µg/mL + modifying enzyme inhibitor 61.44 µg/mL), streptomycin (170 µg/mL + modifying enzyme inhibitor 61.44 µg/mL), and streptomycin (470 µg/mL + modifying enzyme inhibitor 61.44 µg/mL) exhibited 99% viability and less than 5% cytotoxicity in the HSF cell line. No undesirable morphological manifestations, such as cell rounding, shrinkage, or granulation, were observed compared with the negative control. These findings confirm that all tested formulations are both highly effective antibacterial agents and safe for human skin applications.
In this study, combining aminoglycosides with aminoglycoside-modifying enzyme inhibitors achieved strong antibacterial activity while maintaining low cytotoxicity toward human fibroblasts. This supports the feasibility of developing adjuvant therapeutic strategies that restore aminoglycoside efficacy without compromising host safety. The reduced cytotoxicity likely results from several complementary mechanisms. First, co-administration of modifying enzyme inhibitors allows for lower aminoglycoside doses to achieve equivalent antimicrobial effects, thereby reducing intracellular drug accumulation and limiting damage to lysosomal and mitochondrial compartments. Second, selective inhibition of bacterial aminoglycoside-modifying enzymes without affecting host cellular enzymes may prevent off-target interactions that typically contribute to cytotoxicity.62,63 Third, human dermal fibroblasts and keratinocytes possess robust antioxidant defenses and efficient endolysosomal repair mechanisms, which mitigate transient aminoglycoside-induced oxidative and structural stress.64
Recent reports have supported the safety of this combined approach. Silver ions inhibit AAC(2′)-Ia, an aminoglycoside-modifying enzyme responsible for plazomicin resistance, without inducing cytotoxicity in mammalian cells.65 Pyrrolidine pentamine derivatives that inhibit AAC(6′)-Ib and restore kanamycin activity with negligible toxicity toward human embryonic kidney cells. These studies align with the present findings, reinforcing that rationally designed aminoglycoside-modifying enzyme inhibitors can potentiate aminoglycoside activity while maintaining biocompatibility.66
Quantitative morphometric analysis demonstrated a significant dose-dependent reduction in cell size (up to 46.7%) accompanied by a marked increase in cellular aggregation (up to 63.4%). The reduction in projected cell area suggests membrane shrinkage and possible cytoplasmic condensation, which are common indicators of antimicrobial-induced stress. The increased aggregation index may reflect membrane surface alterations leading to enhanced cell-cell adhesion or incomplete cell separation following division. Such clustering has been reported in bacterial populations undergoing oxidative or nanoparticle-induced stress, where membrane permeability changes and extracellular polymeric substance (EPS) release may promote aggregation. The strong linear correlation (R² > 0.9) between treatment concentration and both morphometric parameters supports a concentration-dependent structural effect rather than random variability. These quantitative findings strengthen the morphological evidence and provide statistical validation of treatment-induced cellular damage.
Historically, prolonged or high-concentration aminoglycoside exposure, particularly with gentamicin has been associated with lysosomal phospholipidosis and altered lipid metabolism in fibroblasts.67 However, recent evidence indicates that clinically relevant, short-term, or topical exposure is well tolerated.68 Collectively, these results suggest that through dose optimization, controlled exposure, and the inclusion of enzyme inhibitors, aminoglycoside efficacy can be safely harnessed in dermatologic or wound-care applications with minimal host cell injury. Nevertheless, future studies using 3D or ex vivo skin models are necessary to evaluate long-term or cumulative effects and to investigate the roles of oxidative stress, mitochondrial integrity, and apoptotic signaling in determining safety profiles.
Microscopic evaluation alone is not sufficient to conclude cell viability, as optical factors such as focal depth, cell distribution within the field, and slight variations in illumination may create the impression of altered density. Therefore, viability assessment was confirmed using quantitative assays. The absence of membrane disruption, cell debris accumulation, or morphological distortion in the improved high-resolution images supports the conclusion that treatments did not induce cytotoxic effects. Additionally, the lack of statistically significant differences in CFU counts confirms that the treatments are biologically safe under the tested conditions.
Most current antibiotics lack sufficient efficacy to significantly inhibit resistant bacteria, posing a major challenge in the clinical field. Therefore, additive substances such as enzyme inhibitors should be combined with antibiotics in specific ratios to safely and effectively control resistant bacteria. This study introduces a novel formulation consisting of paromomycin (140 or 410 µg/mL) or streptomycin (170 or 470 µg/mL) in combination with a modifying enzyme inhibitor (61.44 µg/mL), which effectively inhibits resistant S. pneumoniae and S. aureus. Interestingly, antagonistic behavior was observed in this work, as P. aeruginosa, a highly virulent pathogen was found to produce an efficient inhibitor that inactivates the resistance enzymes of other Gram-positive pathogens, thereby allowing aminoglycosides to achieve complete inhibition.
ACKNOWLEDGMENTS
The author is thankful to the Deanship of Scientific Research, Shaqra University, for supporting this work.
FUNDING
None.
DATA AVAILABILITY
All datasets generated or analyzed during this study are included in the manuscript.
ETHICS STATEMENT
Not applicable.
- Zapun A, Contreras-Martel C, Vernet T. Penicillin-binding proteins and β-lactam resistance. FEMS Microbiol Rev. 2008;32(2):361-385.
Crossref - Yadegar A, Sattari M, Mozafari NA, Goudarzi GR. Prevalence of the genes encoding aminoglycoside-modifying enzymes and methicillin resistance among clinical isolates of Staphylococcus aureus in Tehran, Iran. Microb Drug Resist. 2009;15(2):109-113.
Crossref - Eleazar CI, Udoh IP, Maduakor U, Emenuga V, Nweke RN. Streptococcus pneumoniae nasopharyngeal colonization and antibiogram of isolates from paediatric population in Enugu, South East Nigeria. Int J Pathog Res. 2024;13(3):6-14.
Crossref - Anand U, Reddy B, Singh VK, et al. Potential environmental and human health risks caused by antibiotic-resistant bacteria (ARB), antibiotic resistance genes (ARGs) and emerging contaminants (ECs) from municipal solid waste (MSW) landfill. Antibiotics. 2021;10(4):374.
Crossref - Laxminarayan R, Chaudhury RR. Antibiotic resistance in India: drivers and opportunities for action. PLoS Med. 2016;13(3):e1001974.
Crossref - Baltić B, Starčević M, Đorđević J, Mrdović B, Marković R. Importance of medium chain fatty acids in animal nutrition. IOP Conf Ser: Earth Environ Sci. 2017;85(1):012048.
Crossref - Shao Y, Zhen W, Guo F, et al. Pretreatment with probiotics Enterococcus faecium NCIMB 11181 attenuated Salmonella Typhimurium-induced gut injury through modulating intestinal microbiome and immune responses with barrier function in broiler chickens. J Anim Sci Biotechnol. 2022;13(1):130.
Crossref - Kerek A, Csanady P, Jerzsele A. Antibacterial efficiency of propolis—Part 1. Magyar. Allatorvosok Lapja. 2022;144(5):285-298.
- Omonijo FA, Ni L, Gong J, et al. Essential oils as alternatives to antibiotics in swine production. Anim Nutr. 2018;4(2):126-136.
Crossref - Al-Kahtani SN, Alaqil AA, Abbas AO. Modulation of antioxidant defense, immune response, and growth performance by inclusion of propolis and bee pollen into broiler diets. Animals. 2022;12(13):1658.
Crossref - Petrilla J, Matis G, Molnar A, et al. In vitro investigation of the antibacterial efficacy of butyrate on various Campylobacter jejuni strains. Magyar Allatorvosok Lapja. 2021;143(1):57-64.
- Mahlapuu M, Håkansson J, Ringstad L, Björn C. Antimicrobial peptides: An emerging category of therapeutic agents. Front Cell Infect Microbiol. 2016;6:194.
Crossref - Halder N, Joardar SN, Sunder J, et al. Impact of multi-strain probiotics supplementation on growth, immune responses and physiological traits in backyard poultry of Andaman and Nicobar Islands, India. Front Microbiol. 2025;16:1625167.
Crossref - Such N, Molnar A, Pal L, et al. The effect of pre- and probiotic treatment on the Gumboro-titer values of broilers. Magyar Allatorvosok Lapja. 2021;143(2):119-127.
- Abd El-Ghany WA. Paraprobiotics and postbiotics: contemporary and promising natural antibiotic alternatives and their applications in the poultry field. Open Vet J. 2020;10(3):323-330.
Crossref - Heaton CJ, Gerbig GR, Sensius LD, Patel V, Smith TC. Staphylococcus aureus epidemiology in wildlife: a systematic review. Antibiotics. 2020;9(2):89.
Crossref - Chen H, Zhang J, He Y, et al. Exploring the role of Staphylococcus aureus in inflammatory diseases. Toxins. 2022;14(7):464.
Crossref - Abdullahi IN, Zarazaga M, Campana-Burguet A, Eguizabal P, Lozano C, Torres C. Nasal Staphylococcus aureus and S. pseudintermedius carriage in healthy dogs and cats: a systematic review of their antibiotic resistance, virulence and genetic lineages of zoonotic relevance. J Appl Microbiol. 2022;133(6):3368-3390.
Crossref - Bourely C, Cazeau G, Jarrige N, et al. Co-resistance to amoxicillin and tetracycline as an indicator of multidrug resistance in Escherichia coli isolates from animals. Front Microbiol. 2019;10:2288.
Crossref - Dahms C, Huebner NO, Kossow A, Mellmann A, Dittmann K, Kramer A. Occurrence of ESBL-producing Escherichia coli in livestock and farm workers in Mecklenburg-Western Pomerania, Germany. PLoS One. 2015;10(11):e0143326.
Crossref - Pyzik E, Marek A, Stepien-Pysniak D, Urban-Chmiel R, Jarosz LS, Jagielo-Podebska I. Detection of antibiotic resistance and classical enterotoxin genes in coagulase-negative Staphylococci isolated from poultry in Poland. J Vet Res. 2019;63(2):183-190.
Crossref - Boost MV, Wong A, Ho J, et al. Isolation of methicillin-resistant Staphylococcus aureus (MRSA) from retail meats in Hong Kong. Foodborne Pathog Dis. 2013;10(8):705-710.
Crossref - Cha SY, Kang M, Yoon RH, Park CK, Moon OK, Jang HK. Prevalence and antimicrobial susceptibility of Salmonella isolates in Pekin ducks from South Korea. Comp Immunol Microbiol Infect Dis. 2013;36(5):473-479.
Crossref - Zhao F, He S, Tan A, et al. Isolation, identification and character analysis of Streptococcus dysgalactiae from Megalobrama terminalis. J Fish Dis. 2020;43(2):239-252.
Crossref - Dewachter L, Denereaz J, Liu X, et al. Amoxicillin-resistant Streptococcus pneumoniae can be resensitized by targeting the mevalonate pathway as indicated by sCRilecs-Seq. ELife. 2022;11:e75607.
Crossref - Clinical and Laboratory Standards Institute (CLSI). Methods for Antimicrobial Susceptibility Testing of Infrequently Isolated or Fastidious Bacteria Isolated from Animals, 1st ed. CLSI Standards VET06. Wayne, PA: CLSI; 2017.
- Clinical and Laboratory Standards Institute (CLSI). Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically, 12th ed. CLSI standards M07. Wayne, PA: CLSI; 2024.
- Da Silva Filho LVF, Levi JE, Bento CNO, Da Silva Ramos SRT, Rozov T. PCR identification of Pseudomonas aeruginosa and direct detection in clinical samples from cystic fibrosis patients. J Med Microbiol. 1999;48(4):357-361.
Crossref - Seemann T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics. 2014;30(14):2068-2069.
Crossref - Vanderzant C, Don S, eds. Compendium of Methods for the Microbiological Examination of Foods. 3rd ed. American Public Health Association; 1992:331-340
- Lowry OH, Rosebrough NJ, Farr AL, Randall RJ. Protein measurement with the Folin phenol reagent. J Biol Chem. 1951;193(1):265-275.
Crossref - Andrews P. Estimation of the molecular weights of proteins by Sephadex gel-filtration. Biochem J. 1969;91(1):222-233.
Crossref - Blackshear PJ. Systems for polyacrylamide gel electrophoresis. Methods Enzymol. 1984;104:237-255.
Crossref - Laemmli UK. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature. 1970;227(5259):680-685.
Crossref - Fairbanks G, Steck TL, Wallach DFH. Electrophoretic analysis of the major polypeptides of the human erythrocyte membrane. Biochemistry. 1971;10(13):2606-2617.
Crossref - Skehan P, Storeng R, Scudiero D, et al. New colorimetric cytotoxicity assay for anticancer-drug screening. J Natl Cancer Inst. 1990;82(13):1107-1112.
Crossref - Allam RM, Al-Abd AM, Khedr A, et al. Fingolimod interrupts the cross-talk between estrogen metabolism and sphingolipid metabolism within prostate cancer cells. Toxicol Lett. 2018;291:77-85.
Crossref - Kong J, Wu ZX, Wei L, Chen ZS, Yoganathan S. Exploration of antibiotic activity of aminoglycosides, in particular ribostamycin alone and in combination with ethylenediaminetetraacetic acid against pathogenic bacteria. Front Microbiol. 2020;11:1718.
Crossref - Shankar N, Lockatell CV, Baghdayan AS, Drachenberg C, Gilmore MS, Johnson DE. Role of Enterococcus faecalis surface protein Esp in the pathogenesis of ascending urinary tract infection. Infect Immun. 2001;69(7):4366-4372.
Crossref - Baraboutis IG, Tsagalou EP, Lepinski JL, et al. Primary Staphylococcus aureus urinary tract infection: the role of undetected hematogenous seeding of the urinary tract. Eur J Clin Microbiol Infect Dis. 2010;29(9):1095-1101.
Crossref - Alaa MA, Ramadan H, Eman SA. The effect of EDTA in combination with some antibiotics against clinical isolates of Gram-negative bacteria in Mansoura, Egypt. Microb Pathog. 2021;154:104840.
Crossref - Martins VV, Macarenco AC, Gradella DG, Stehling EG. Antagonism between clinical and environmental isolates of Pseudomonas aeruginosa against coliforms. Water Supply. 2014;14(1):99-106.
Crossref - Vasconcelos U, Calazans GMT, Andrade MAG, Medeiros LV. Evidence of the antagonism between Pseudomonas aeruginosa and indicating bacteria of fecal contamination in water. [Evidência do antagonismo entre Pseudomonas aeruginosa e bactérias indicadoras de contaminação fecal em água]. Higiene Alimentar. 2006;20(140):127-130.
- Daniel JF, Jeffery KK, Douglas LW. Antagonism of foodborne bacteria by Pseudomonas spp.: a possible role for iron. J Food Prot. 1989;52(7):484-489.
Crossref - McCullers KW, McCullers JE, Chang AT, et al. Streptococcus pneumoniae and Pseudomonas aeruginosa pneumonia induce distinct host responses. Crit Care Med. 2010;38(1):223-241.
Crossref - Nguyen AT, Oglesby-Sherrouse A. Interactions between Pseudomonas aeruginosa and Staphylococcus aureus during co-cultivations and polymicrobial infections. Appl Microbiol Biotechnol. 2016;100(14):6141-6148.
Crossref - Jama-Kmiecik A, Maczynska B, Frej-Madrzak M, et al. The changes in the antibiotic resistance of Staphylococcus aureus, Streptococcus pneumoniae, Enterococcus faecalis and Enterococcus faecium in the clinical isolates of a multiprofile hospital over 6 years (2017–2022). J Clin Med. 2025;14(2):332.
Crossref - Kakoullis L, Papachristodoulou E, Chra P, Panos G. Mechanisms of antibiotic resistance in important Gram-positive and Gram-negative pathogens and novel antibiotic solutions. Antibiotics. 2021;10(4):415.
Crossref - Gergova R, Boyanov V, Muhtarova A, Alexandrova A. A review of the impact of streptococcal infections and antimicrobial resistance on human health. Antibiotics. 2024;13(3):360.
Crossref - El-Gammal MS, Abouwarda AM, Abdulkhair WM. In vitro inhibition of aminoglycoside-resistant Staphylococcus aureus by modifying enzyme inhibitory protein of Pseudomonas aeruginosa. Afr J Microbiol Res. 2013;7(25):3262-3268.
Crossref - Culebras E, Martinez JL, Baquero F, Perez-Diaz JC. pH modulation of aminoglycoside resistance in Staphylococcus epidermidis harbouring 62 -N-aminoglycoside acetyltransferases. J Antimicrob Chemother. 1996;37(5):881-889.
Crossref - Jouybari MA, Ahanjan M, Mirzaei B, Goli HR. Role of aminoglycoside-modifying enzymes and 16S rRNA methylase (ArmA) in resistance of Acinetobacter baumannii clinical isolates against aminoglycosides. Rev Soc Bras Med Trop. 2021;54(5):e05992020.
Crossref - Cui G, Pan C, Xu P, et al. Purification and characterization of a novel bacteriocin produced by Enterococcus faecalis CG-9 from human saliva. Biotechnol Biotechnol Equip. 2020;34(1):1224-1233.
Crossref - Chen C, Chen X, Jiang M, Rui X, Li W, Dong M. A newly discovered bacteriocin from Weissella hellenica D1501 associated with Chinese Dong fermented meat (Nanx Wudl). Food Control. 2014;42:116-124.
Crossref - Liu H, Zhang L, Yi H, Han X, Chi C. Identification and characterization of plantaricin Q7, a novel plantaricin produced by Lactobacillus plantarum Q7. LWT Food Sci Techno. 2016;71:386-390.
Crossref - Krause KM, Serio AW, Kane TR, Connolly LE. Aminoglycosides: an overview. Cold Spring Harb Perspect Med. 2016;6(6):a027029.
Crossref - Kohanski MA, Dwyer DJ, Hayete B, Lawrence CA, Collins JJ. A common mechanism of cellular death induced by bactericidal antibiotics. Cell. 2007;130(5):797-810.
Crossref - Pérez-Varela M, Corral J, Aranda J, Barbé J. Functional Characterization of AbaQ, a Novel Efflux Pump Mediating Quinolone Resistance in Acinetobacter baumannii. Antimicrob Agents Chemother. 2018;62(9):e00906-18.
Crossref - Nizet V, Ohtake T, Lauth X, et al. Innate antimicrobial peptide protects the skin from invasive bacterial infection. Nature. 2001;414(6862):454-457.
Crossref - Krause KM, Serio AW, Kane TR, Connolly LE. Aminoglycosides: An Overview. Cold Spring Harb Perspect Med. 2016;6(6):a027029.
Crossref - Daigle DM, Hughes DW, Wright GD. Prodigious substrate specificity of AAC(6′)-APH(22′), an aminoglycoside resistance determinant in enterococci and staphylococci. Chem Biol. 1999;6(2):99 110.
Crossref - Garneau Tsodikova S, Labby KJ. Mechanisms of resistance to aminoglycoside antibiotics: overview and perspectives. MedChemComm. 2016;7(1):11 27.
Crossref - Angel JM, Jan S, Clemencia P, et al. Restoring susceptibility to aminoglycosides: identifying small molecule inhibitors of enzymatic inactivation. RSC Med Chem. 2023;14(9):1591–1602.
Crossref - Dumont C, Wuestenberghs F, Lanthier N, Piessevaux H, Dahlqvist G. Malnutrition is highly prevalent in hospitalized cirrhotic patients and associates with a poor outcome. Acta Gastro-Enterologica Belgica. 2022;85(2): 311-319.
Crossref - Ngo D, Magana AJ, Tran T, et al. Inhibition of Enzymatic Acetylation-Mediated Resistance to Plazomicin by Silver Ions. Pharmaceuticals. 2023;16(2):236.
Crossref - Sklenicka J, Tran T, Ramirez MS, et al. Structure–Activity Relationship of Pyrrolidine Pentamine Derivatives as Inhibitors of the aminoglycoside 62 -N-acetyltransferase type Ib. Antibiotics. 2024;13(7):672.
Crossref - Regec AL, Trump BF, Trifillis AL. Effect of gentamicin on the lysosomal system of cultured human proximal tubular cells. Endocytotic activity, lysosomal pH and membrane fragility. Biochem Pharmacol. 1989;38(15):2527-2534.
Crossref - Rosenberg CR, Fang X, Allison KR. Potentiating aminoglycoside antibiotics to reduce their toxic side effects. PLoS One. 2020;15(9):e0237948.
Crossref
© The Author(s) 2026. Open Access. This article is distributed under the terms of the Creative Commons Attribution 4.0 International License which permits unrestricted use, sharing, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.