


ISSN: 0973-7510
E-ISSN: 2581-690X
The gut microbiome plays a critical role in heart failure (HF) through immune, metabolic, and inflammatory pathways. Altered microbial composition and metabolites may contribute to HF pathogenesis and progression. This systematic review compared gut microbiome profiles in adults with HF and healthy controls (HCs), highlighted microbial taxa linked to HF incidence, and examined functional and metabolic pathway changes. Eligible studies included peer-reviewed research comparing gut microbiota in adults with HF and HCs reporting taxonomic, functional, or metabolite outcomes. A systematic search of PubMed, Scopus, and Web of Science identified studies published between January 2021 and August 2025. Data extraction used a structured table to capture study characteristics, while quality was assessed with the Newcastle–Ottawa Scale. Ten studies involving 407 HF patients and 327 HCs were reviewed. Heart failure patients showed reduced microbial richness and diversity compared with healthy controls. Depletion of beneficial short-chain fatty acid (SCFA) producing taxa such as Firmicutes (Faecalibacterium, Ruminococcaceae, Lachnospiraceae) and Actinobacteria (Bifidobacterium) was common, while enrichment of opportunistic Proteobacteria (Klebsiella, Escherichia, Enterobacteriaceae) was reported. Functional analyses highlighted downregulation of butyrate- and propionate-producing pathways and increased pro-inflammatory metabolites production, particularly trimethylamine N-oxide (TMAO). Subtype differences were suggested between HFrEF and HFpEF, though HFmrEF was not examined. Limitations included methodological heterogeneity, lack of longitudinal designs, and restriction to English-language studies. HF is consistently associated with gut dysbiosis, marked by loss of SCFA-producing commensals and enrichment of pathogenic taxa with harmful metabolic potential. Broader, longitudinal, and subtype-specific studies required to establish causality and guide microbiota-targeted therapies in HF.
Gut Microbiota, Dysbiosis, Heart Failure, Firmicutes, Proteobacteria, TMAO
Heart failure (HF) is a complex and progressive clinical syndrome characterized by impaired ventricular filling and/or pumping capacity, or by structural and functional abnormalities that lead to inadequate cardiac output or increased filling pressures.1 It affects more than 64 million people worldwide, representing a major contributor to morbidity, mortality, and healthcare burden.2 HF is categorized based on left ventricular ejection fraction (EF) into reduced EF (HFrEF, ≤40%), mildly reduced EF (HFmrEF, 41%-49%), and preserved EF (HFpEF, ≥50%).3 Despite advancements in diagnostic tools, pharmacological agents, and device-based therapies, HF continues to impose a substantial socioeconomic burden, with recurrent hospitalizations, reduced productivity, and long-term care requirements. Current treatments alleviate symptoms and improve survival in certain patient subgroups but fail to halt disease progression or fully restore cardiac function, highlighting the need to explore novel mechanistic pathways.4
The human gut microbiota composed of trillions of microorganisms including bacteria, archaea, fungi, and viruses, plays a critical role in maintaining host homeostasis through nutrient metabolism, immune regulation, and preservation of gut barrier integrity.5 Disruption of this microbial ecosystem, termed dysbiosis, has been implicated in a wide range of diseases, including gastrointestinal, metabolic, neurologic, and cardiovascular disorders.6 Through the production of bioactive metabolites such as trimethylamine N-oxide (TMAO), short-chain fatty acids (SCFAs), and bile acids (BAs), the gut microbiota exerts systemic effects on host metabolism, inflammation, endothelial function, and cardiovascular regulation which this interaction often referred to as the “gut-heart axis”.7
Emerging evidence suggests that gut microbiota dysbiosis contributes to the pathogenesis of several cardiovascular conditions, including hypertension, atherosclerosis, and myocardial infarction.8 Elevated TMAO levels have been linked to endothelial dysfunction and adverse cardiovascular outcomes, whereas reduced SCFA production may impair vascular homeostasis and energy balance.7 In HF, dysbiosis is thought to result from intestinal congestion, reduced perfusion, and altered BAs metabolism, which collectively compromise gut barrier integrity and increase intestinal permeability. This facilitates the translocation of bacteria and endotoxins such as lipopolysaccharides (LPS) into systemic circulation, triggering low-grade inflammation and immune dysregulation.9
Several studies have reported that HF patients exhibit reduced microbial diversity, depletion of beneficial taxa (e.g., Ruminococcaceae, Lachnospiraceae, Faecalibacterium), and enrichment of pathogenic species (e.g., Enterococcus, Escherichia, Klebsiella).10 These alterations modify key metabolites, including SCFAs, TMAO, BAs, and amino acids, which play central roles in modulating inflammation, endothelial function, and cardiovascular outcomes. Consequently, the gut-heart axis represents a dynamic interplay where impaired cardiac function exacerbates gut dysfunction, and microbial/metabolic disturbances accelerate HF progression.11
Rationale
Despite the growing body of research, current evidence remains fragmented, with many studies focusing on either taxonomic alterations or metabolite pathways in isolation, without providing an integrated
perspective.12,13 Importantly, a lack of systematic review has comprehensively synthesized evidence from the past five years (2021-2025) on gut microbial composition, functional alterations, and their clinical implications in HF. Given the increasing recognition of the gut-heart axis as a potential therapeutic target, a rigorous and up-to-date synthesis is essential to inform future research and guide the development of microbiota-based interventions.
Objectives
This systematic review aims to critically evaluate and synthesize the most recent evidence on gut microbiota alterations in adults with HF. Specifically, it seeks to:
- Compare the gut microbiome composition between HF patients and HCs;
- Determine gut microbial taxa associated with the incidence of HF; and
- Examine alterations in functional and metabolic pathways of the gut microbiome in HF.
By integrating compositional and functional evidence, this review provides a comprehensive perspective on the gut-heart axis in HF, highlighting its potential as a diagnostic, prognostic, and therapeutic target. The findings may inform the design of future clinical trials and foster the development of dietary, probiotic, or microbiota-targeted therapies aimed at improving outcomes in HF patients.
Protocol registration and PRISMA guideline
A systematic review protocol was developed and prospectively registered with PROSPERO (CRD420251145733). The writing of this systematic review was compliant with the 2020 Preferred Reporting Items for Systematic Reviews and Meta-analyses (PRISMA) guideline (Supplementary Tables S1 and S2).
Identification and selection of papers
This systematic review was conducted in accordance with the PRISMA guidelines to ensure methodological rigor, transparency, and reproducibility. The review process comprised defining research questions, a comprehensive literature search, application of predefined eligibility criteria, selection of studies, data extraction, and synthesis of findings.
Research questions
- How does the gut microbiome composition of adults with HF differ from that of HCs in studies published between 2021 and 2025?
- What gut microbial taxa have been consistently reported to be associated with the incidence of HF?
- What alterations in functional and metabolic pathways of the gut microbiome are observed in HF patients compared with HCs?
Eligibility criteria
Studies were included or excluded based on predefined criteria outlined in Table 1. These criteria specified the study population, design, outcomes, publication, timeframe, and language requirements, ensuring that only relevant research was selected for this review.
Information sources/databases
A systematic search of peer-reviewed literature was performed on 18 August 2025 across three electronic databases: PubMed, Scopus, and Web of Science, from database inception to the search date. The search combined Medical Subject Headings (MeSH) and free-text keywords related to “gut microbiota” and “heart failure”, along with their synonyms. The full search strategies for each database are provided in Table 2.
Table (1): Major Compounds identified in the chloroform and ethanol extract of F. indica using GC-MS
Inclusion Criteria |
Exclusion Criteria |
|---|---|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Search strategy
Database-specific strategies were developed using a combination of controlled vocabulary (MeSH) and free-text terms. Boolean operators (“AND”, “OR”) were used to combine search concepts, and filters were applied to limit results to human studies, adults (≥18 years), and publications between 2021 and 2025. Table 2 presents the complete search strings, including keywords, MeSH terms, applied filters, for PubMed, Scopus, and Web of Science and date of access.
Table (2): Search Strategy for Each Database
Database |
Search strategy |
Date of access |
|---|---|---|
SCOPUS |
TITLE-ABS-KEY (“heart failure” OR HFrEF OR HFpEF) AND TITLE-ABS-KEY (“gut microbiota” OR “gut microbiome” OR “intestinal microbiota”) AND (LIMIT-TO (LANGUAGE, “English”)) AND (LIMIT-TO (DOCTYPE, “ar”)) AND (PUBYEAR > 2020 AND PUBYEAR
| 12/8/2025 |
PubMed |
(“Heart Failure”[MeSH] OR “Heart Failure”[tiab] OR “Congestive Heart Failure”[tiab] OR “Cardiac Failure”[tiab]) AND (“Gastrointestinal Microbiome”[MeSH] OR “Gut Microbiome”[tiab] OR “Gut Microbiota”[tiab] OR “Intestinal Microbiome”[tiab] OR “Intestinal Microbiota”[tiab] OR “Microbiota”[tiab] OR “Microbiome”[tiab]) |
13/8/2025 |
Web of Science |
TS= (“heart failure” OR HFpEF OR HFrEF) AND TS= (“gut microbiota” OR “gut microbiome” OR “intestinal microbiota”) AND LA = (English) AND DT= (Article) AND PY= (2021-2025) |
13/8/2025 |
Study selection process/screening
All search results obtained from each online database were downloaded in CSV format. The CSV files were merged into a single master file, and duplicate entries were removed. Two independent reviewers then screened the titles and abstracts of the retrieved articles according to the predefined eligibility criteria. The full texts of potentially eligible studies were subsequently assessed for inclusion. Any disagreements between reviewers were resolved through discussion or, when necessary, by consultation with a third reviewer. The study selection process followed the PRISMA 2020 flow diagram, documenting the number of records identified, screened, excluded (with reasons), and included in the final review.
Data extraction
Data from the included studies were collected using a standardized extraction form, capturing the following details as shown in Table 3.
Table (3): Data Extraction Framework for Included Studies
Category |
Details |
|---|---|
Study details |
First author, Year, Country, Journal |
Study design |
Study design (RCT, Cohort, Case-control, Cross-sectional), Study period |
Population |
Sample size, Age, Gender, Case subtype, Comorbidities, New York Heart Association classification |
Comparison |
Control group (healthy controls), Heart failure patient |
Methodology |
Sequencing method |
Exposure/Characteristics |
Medications consumed, Lifestyle |
Quality assessment
The Newcastle-Ottawa Scale (NOS) is widely applied for quality assessment of non-randomized studies. The tool evaluates three broad domains: Selection, Comparability, and Outcome. For cohort studies, the NOS awards a maximum of (4) points for Selection, (2) points for Comparability, and (3) points for Outcome, giving a total score of (9). In contrast, when adapted for cross-sectional studies, the NOS allocates (5) points to Selection, (2) points to Comparability, and (2) points to Outcome, also resulting in a total maximum score of (9). The overall NOS scores were classified into three categories: low/poor quality (0-3), moderate quality (4-6), and high quality (7-9).
Study selection
The database search initially retrieved 570 studies, of which 10 duplicates were removed. Following the title and abstract screening of 560 studies, 78 full-text articles were reviewed against the eligibility criteria. A total of ten studies were included in this systematic review; the reasons for excluding the full-text articles are provided in Figure 1.
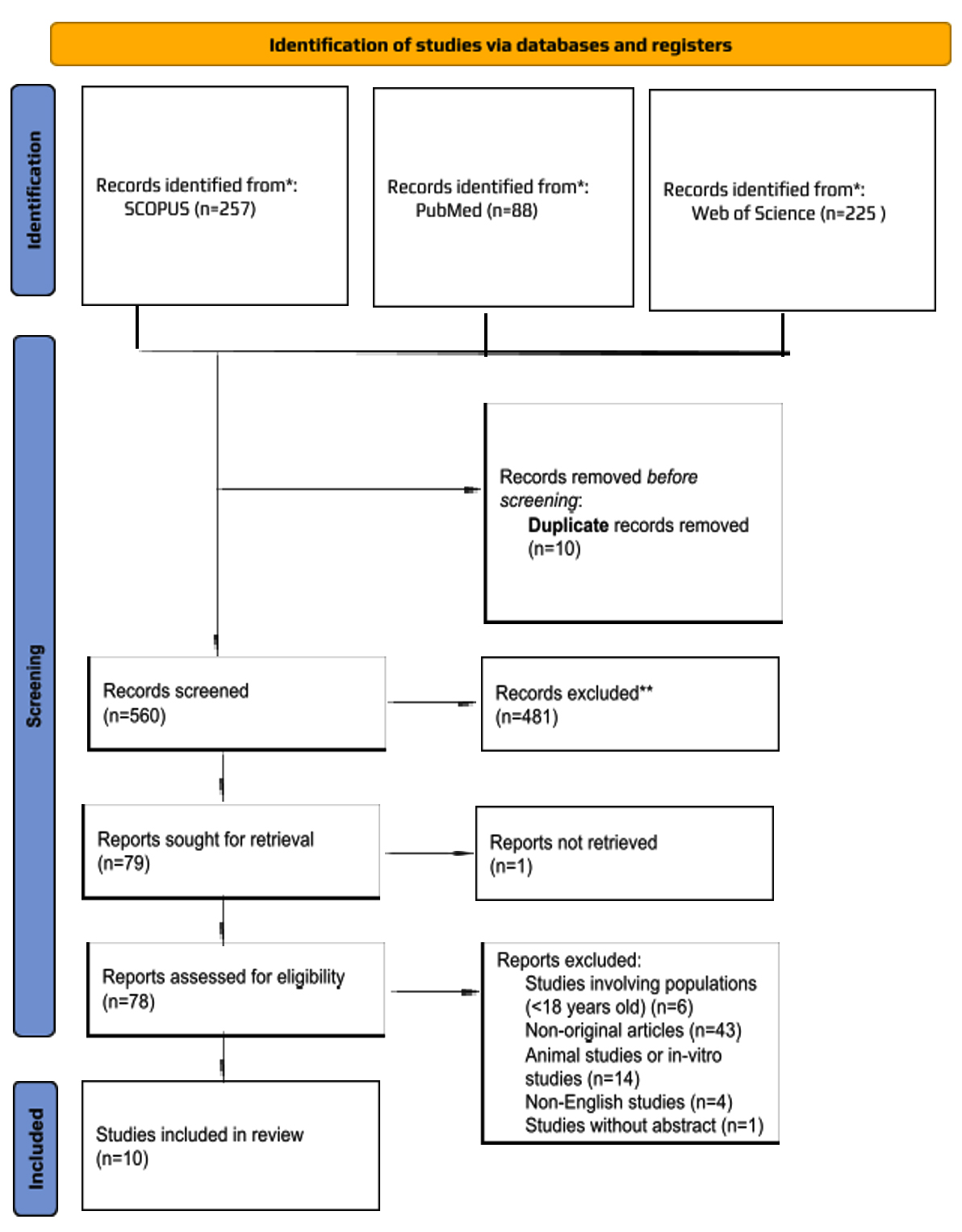
Figure 1. Flow Diagram of The Proposed Searching Study
Study characteristics
A total of ten observational studies involving 734 participants (407 HF patients and 327 HCs) were included, published between 2021 and 2025 as shown in Supplementary Table S3. Eight studies employed a cross-sectional design14-21 and two used a cohort design.22,23 The studies were conducted in China (n = 6), Australia (n = 2), Russia (n = 1), and Japan (n = 1). Most HF patients were classified as NYHA class II-IV.
Regarding HF subtypes, one study focused solely on HFrEF,19 one on HFpEF,22 three included mixed HF subtypes (HFrEF, HFpEF, HFmrEF)17,20,23 and the remaining studies did not specify the HF subtype. Common comorbidities reported included hypertension, atrial fibrillation, and diabetes.14,15,17,19-21,23
All studies utilized 16S rRNA sequencing, amplifying the V3-V4 region. One study used 16S rDNA sequencing18 and two used alternative methods: gene amplicon analysis23 and metagenomic sequencing.15
Medication reporting varied. Most studies documented beta-blocker use among HF patients, whereas three15,18,22 provided no detailed medication data. Similarly, smoking and alcohol status were not reported in two studies.19,22
The extent of confounder adjustment differed notably across studies. Some cross-sectional studies did not adjust for any confounders,19 limiting the reliability of their associations. In contrast,14,17,20 others implemented comprehensive adjustments for key factors such as age, gender, lifestyle, comorbidities, and medications. Study by Huang et al.,15 partially controlled for confounders (age, gender, BMI, smoking, alcohol) and excluded participants with antibiotics, probiotics, or major comorbidities. Zhang et al.,21 adjusted for age, gender, and cardiovascular disease,21 while some studies16,18 reported no clear adjustments. Among the cohort studies,22 accounted for BMI, age, hypertension, diet and gender, while Emoto et al.23 further adjusted for age, gender, and comorbidities, enhancing robustness.
Quality assessment
The overall quality of the included studies ranged from moderate to high based on the Newcastle-Ottawa Scale (NOS), with scores between 5 and 8 (Supplementary Tables S4-S6). Six studies were rated high quality (NOS 7-8), while four were of moderate quality (NOS 5-6).
High-quality studies generally applied robust methodologies, including validated sequencing and metabolomic platforms, independent outcome assessment, and adjustment for major confounders (age, gender, comorbidities, medications, and in some cases diet or BMI). Notable examples include14,15,17,20,21 which all controlled for multiple risk factors and applied advanced analytic pipelines. These studies achieved NOS scores of 7-8, reflecting low risk of bias.
In contrast, moderate-quality studies, such as16,18,19 scored lower (5-6) mainly due to limited adjustment for confounders, small sample sizes, or incomplete reporting of recruitment and response rates. While they employed validated sequencing and statistical methods, the absence of confounder control reduced internal validity.
The two cohort studies also varied in quality. Study by Beale et al.,22 scored 6/9, reflecting moderate quality, as it adjusted for key confounders but lacked follow-up data typical of stronger cohort designs.22 One of the included studies, scored 7/9, achieving high quality due to comprehensive adjustment and appropriate microbiome analyses, despite being single center.23
Gut microbiota richness and diversity (Alpha Diversity) in HF patients compared with HCs
A total of nine studies assessed alpha diversity to compare gut microbiota richness and/or diversity between patients with HF and HCs as shown in Table 4. One study reported only microbial diversity, whereas the remainder evaluated both richness and diversity. Across the included studies, a range of alpha diversity indices were employed, including Shannon, Simpson, Chao1, ACE, Faith’s phylogenetic diversity (PD)-whole-tree, Pielou’s evenness, and observed operational taxonomic units (OTUs).
Table (4): Summary of the Gut Microbiota Alpha and Beta Diversity Results in HF Patients
| Studies | Alpha Diversity | Beta Diversity | |||||
|---|---|---|---|---|---|---|---|
| Indices/Metrics used | Microbial Richness | Microbial Diversity | Significant tests | Indices/Metrics used | Significant | Significant tests | |
| HFrEF | |||||||
| Yafarova et al.19 | Pielou, Chao1, PD-whole-tree | ↑# | ↑# | Mann-Whitney/χ2, Kruskal-Wallis | Euclidean (Manhattan, Minkowski), Canberra, Bray-Curtis, Jaccard | Significantly different gut microbiota composition between HF patients and control group | PERMANOVA |
| HFpEF | |||||||
| Beale et al.22 | OTUs, Shannon, Chao1 | ↓ | ↓ | Student’s t-test | Jaccard, Bray-Curtis, unweighted, weighted UniFrac | Significantly different gut microbiota composition between HF patients and control group | PERMANOVA |
| HFrEF + HFpEF + HFmrEF | |||||||
| Yang et al.20 | Chao1, ACE, Shannon, Simpson | ↓ | ↓ | ANOVA, t-test, Kruskal-Wallis, Mann-Whitney, χ2 | UniFrac | Significantly different gut microbiota composition between HF patients and control group | ANOVA, t-test, Kruskal-Wallis, Mann-Whitney, χ2 |
| Sun et al.17 | Chao1, PD-whole-tree, Shannon | ↓# | ↓# | Welch’s t-test method | UniFrac | Significantly different gut microbiota composition between HF patients and control group | Welch’s t-test method |
| Emoto et al.23 | NR | NR | NR | NR | NR | NR | NR |
| Unspecified CHF | |||||||
| Ahmad et al.14 | Shannon | NR | ↓ | Mann-Whitney, Wilcoxon matched pairs signed rank, Spearman’s rank tests | NR | NR | NR |
| Huang et al.15 | Chao, Shannon, Simpson | ↓ | ↓ | Wilcoxon rank sum test | Unweighted UniFrac | Significantly different gut microbiota composition between HF patients and control group | Wilcoxon rank sum test |
| Zhang et al.21 | Shannon, Chao1 | ↓# | (=) | Independent samples t-test, one-way ANOVA, Mann-Whitney, Kruskal-Wallis test | Non-metric multidimensional scaling | Significantly different gut microbiota composition between HF patients and control group | RDA (Redundancy Analysis) and CCA (Canonical Correspondence Analysis) |
| Peng et al.16 | Observed, Chao1, Shannon, Simpson | ↓# | ↓# | Kruskal-Wallis test | Bray-Curtis | Significantly different gut microbiota composition between HF patients and control group | Adonis test |
| Wang et al.18 | Chao1 | ↓# | ↓# | Wilcoxon rank sum test, unpaired two-tailed, Student’s t-test | Bray–Curtis, Weighted UniFrac | Significantly different gut microbiota composition between HF patients and control group | Wilcoxon rank sum test |
#: Significant difference; (=) no change/similar; ↑ increased; ↓ decreased; CHF: chronic heart failure; HF: heart failure; HFmrEF: heart failure with mildly reduced ejection fraction; HFpEF: heart failure with preserved ejection fraction; HFrEF: heart failure with reduced ejection fraction; NR: not reported; OTUs: observed taxonomic units; PD-whole tree: phylogenetic diversity (whole tree).
Overall, most studies consistently demonstrated a reduction in microbial richness and diversity among HF patients compared with HCs, regardless of the specific indices used. However, heterogeneity was observed across HF subtypes. In particular, one study on HFrEF patients reported increased microbial richness and diversity relative to controls 19 which contrasts with the overall trend. For HFpEF, the available evidence indicated reduced richness and diversity.22 Studies that combined multiple HF phenotypes (HFrEF, HFmrEF, and HFpEF) consistently showed lower richness and diversity compared with HCs.17,20
For studies involving patients with unspecified chronic HF, findings were less uniform. While most reported reduced microbial richness and diversity15,16,18 one study found no significant difference in microbial diversity despite reduced richness.21 Another study reported a reduction in diversity without assessing richness.14
Taken together, these findings suggest that HF is generally associated with decreased gut microbiota richness and diversity, although exceptions exist, particularly in HFrEF populations. The variability in results may reflect differences in HF subtypes, patient characteristics, and methodological approaches, including the choice of sequencing platforms and diversity indices.
Gut Microbiota Composition (Beta Diversity) in HF patients compared with HCs
Of the ten studies,14,23 did not report the beta diversity analysis in HF patients against HCs. The beta diversity, which explored the similarity of gut microbiota composition between the comparison groups, was assessed using the Bray–Curtis, Jaccard, Canberra, Euclidean, and both weighted and unweighted UniFrac. Ordination approaches such as non-metric multidimensional scaling and principal coordinate analysis were commonly used.
All the studies that reported the beta diversity outcomes demonstrated a significantly different gut microbiota composition between patients with HF and HCs across all spectrums of HF, either HFrEF or HFpEF as presented in Table 4.
Change in relative abundance of Gut microbiota taxonomy in HF patients compared with HCs
The summary of changes in the relative abundance of gut microbiota taxonomy in HF patients compared to HCs reported by each study is summarized in Table 5. To summarize, a total of five phyla, nine families, 39 genera, and seven species were reported in the included studies regardless of the HF subtypes. At the phylum level, the relative abundance of Firmicutes was reported in four studies to be lower in HF patients than HCs, and two studies showed a decrease in Bacteroidota in HF patients. Meanwhile, studies have reported increased Proteobacteria, Actinobacteria and Bacillota in HF patients. One study reported an increase in Bacteroidota alone,15 while others showed mixed trends depending on HF subtype.
Table (5): Increased/Decreased/No Change in the Relative Abundance of the Gut Microbiota Taxonomy in HF Patients Compared to HCs
| First author | Phylum | Family | Genus | Species | Assessment method/ Significant tests |
|---|---|---|---|---|---|
| HFrEF | |||||
| Yafarova et al.19 | ↑Bacillota, Actinomycetota# ↓Bacteroidota, Firmicutes# |
↑Acidaminococcaceae# | NR | NR | quasi-Poisson, MaAsLin2 analysis/Spearman test, Mann-Whitney/χ2, Kruskal-Wallis, Shapiro-Wilk test |
| HFpEF | |||||
| Beale et al.22 | NR | NR | ↑Akkermansia#, Sutterella#, Ammoniphilus#, Megasphaera#, Acholeplasma#, Bacteroides#, Erwinia#, Streptococcus# ↓Caldicellulosiruptor#, Ruminococcus#, Mitsuokella# |
NR | QIIME2, DADA2 plugin through q2-DADA2/Student’s t-test, chi-square test |
| HFrEF + HFpEF + HFmrEF | |||||
| Yang et al.20 | ↑Proteobacteria# ↓Firmicutes# |
NR | ↑Enterobacter#, Shigella# ↓Lachnospira#, Parasutterella#, Roseburia#, Barnesiella#, Faecalibacterium#, Dialister# |
NR | LefSe analysis/ANOVA, t-test, Kruskal-Wallis, Mann-Whitney, χ2 |
| Sun et al.17 | ↑Proteobacteria#, | ↓Oscillospiraceae | ↑Shigella#, | ↓Lachnospira | LefSe analysis/ |
| Emoto et al.23 | NR | NR | ↑Klebsiella, Bilophia, Escherichia | NR | Two-tailed Student’s t-test, Mann-Whitney, Spearman’s correlation test |
| Unspecified CHF | |||||
| Ahmad et al.14 | NR | ↓Lachnospiraceae, Oscillospiraceae | ↓Lachnospira | NR | NR/Mann-Whitney, Wilcoxon matched pairs signed rank, Spearman’s rank tests |
| Huang et al.15 | ↑Bacteroidota | NR | ↑Azospirillum#, Prevotella#, Enterocloster#, Pedobacter#, Ruminiclostridium#, Faecalibacterium# | NR | LEfSe/Student’s t-test, Kruskal-Wallis test, Wilcoxon rank sum test, Spearman’s rank correlation. |
| Zhang et al.21 | ↓Firmicute# | ↑Enterobacteriaceae# | ↑Shigella#, Bifidobacterium#, Klebsiella#, Lactobacillus# | NR | LEfSe/Chi-square-test, independent samples t-test, one-way ANOVA, Mann-Whitney U test, Kruskal-Wallis test, Spearman’s rank correlation |
| Peng et al.16 | ↑Proteobacteria, Actinobacteria | ↑Nocardiaceae#, Pseudonocardiaceae#, Leuconostocaceae# | ↑Shigella, Bacteroides, Streptococcus, Akkermansia, Slackia# | NR | LEfSe/Student’s t-test, Mann Whitney U test, Spearman’s rank correlation |
| Wang et al.18 | NR | NR | ↑Shigella#, Lactobacillus#, Atopobium#, Romboutsia#, Streptococcus#, Haemophilus#, Klebsiella#, Campylobacter#, Salmonella# | ↑Yersinia enterocolitica, Candida species, Ruminococcus gnavus group, Ruminococcaceae UCG 005# ↓Eubacterium rectale, Dorea longicatena |
LefSe/Unpaired two-tailed Student’s t-test, Spearman’s rank correlation |
#: Significant difference; ↑ increased; ↓ decreased; CHF: chronic heart failure; HF: heart failure; HFmrEF: heart failure with mildly reduced ejection fraction; HFpEF: heart failure with preserved ejection fraction; HFrEF: heart failure with reduced ejection fraction; NR: not reported.
At the family level, an increased abundance of Acidaminococcaceae, Enterobacteriaceae, Nocardiaceae, Pseudonocardiaceae, and Leuconostocaceae was observed, whereas a decrease in Lachnospiraceae and Oscillospiraceae was reported in HF patients. As for the genus level, there was a greater abundance of Enterobacter, Shigella, Klebsiella, Streptococcus, Akkermansia, Sutterella, Bacteroides, Lactobacillus, Prevotella, Enterocloster, Pedobacter, Ruminiclostridium, Bilophila, Megasphaera, Acholeplasma, Erwinia, Slackia, Nocardiaceae-related genera and others, while a reduced abundance was noted for Faecalibacterium, Lachnospira, Roseburia, Dialister, Barnesiella, Caldicellulosiruptor, and Mitsuokella.
When considering HFrEF specifically, an increased abundance of Bacillota and Actinomycetota was reported, while Bacteroidota and Firmicutes were decreased.19 For HFpEF, only one study provided taxonomic information, reporting multiple genus-level changes without clear phylum-level data.22 In mixed HF phenotypes, increases in Proteobacteria were commonly observed, along with reductions in Firmicutes and Bacteroidota. At the species level, only two studies reported significant findings, with an increase in Yersinia enterocolitica, Candida spp., and Ruminococcus gnavus group, and a decrease in Eubacterium rectale, Dorea longicatena and Lachnospiraceae FCS020.17,18
Changes in Gut-Derived Metabolites and Functional Pathways in HF Patients
Across ten included studies, gut microbiota derived metabolites and associated pathways in HF patients demonstrated a consistent pattern of increased pro-inflammatory metabolites and reduced cardioprotective metabolites as shown in Table 6. TMAO emerged as the most consistently reported metabolite. While one study19 in HFrEF patients observed reduced levels, the majority identified elevated TMAO concentrations, commonly linked to enrichment of Escherichia, Klebsiella, and other TMAO-producing taxa. Elevated TMAO was further associated with adverse ventricular remodeling, greater disease severity, and poor prognosis.
Table (6): Summary of Gut-Derived Metabolites, Functional Pathways and Clinical Implications in HF
| Studies | Metabolites/ Functional pathways |
Associated mechanisms and clinical implications |
Assessment method/ Significant tests |
|---|---|---|---|
| HFrEF | |||
| Yafarova et al.29 | • ↑TMAO# • (=) Indoxyl sulfate |
• NR | • NR/Mann-Whitney |
| HFpEF | |||
| Beale et al.22 | • ↓SCFAs | • Reduce SCFAs production due to decrease in Ruminococcus, leads to development and exacerbation of HFpEF | • NR/Student’s t-test, chi-square test |
| HFrEF + HFpEF + HFmrEF | |||
| Yang et al.20 | • ↑trehalose degradation#, proline degradation#, anaerobic fatty acid β-oxidation#, phenylalanine degradation#, formidoxin oxidoreductase#, arginine degradation III, pentose phosphate pathway (oxidation phase)#, lysine degradation III, asparagine degradation#, glycerol degradation# | • Enterobacter and Shigella modulate lysine degradation metabolism, promoting intestinal inflammation in CHF patients | • GO and KEGG databases, facilitated by PICRUSt2 software, metagenome Seq/ANOVA, t-test, Kruskal-Wallis, Mann-Whitney, χ2 |
| Sun et al.17 | • ↓SCFAs# • ↑lactic acid# |
• Reduction in SCFAs-producing bacteria (Ruminococcaceae, Lachnospiraceae, Dialister) leads to impaired cardioprotective effects • Increased in lactic-acid producing bacteria (Enterococcus, Enterococcaceae) linked to adverse cardiac metabolism |
• PICRUSt software /Welch’s t-test method |
| Emoto et al.23 | • ↑TMAO | • Increase in TMAO due to enrichment of Escherichia or Klebsiella associated with greater HF severity and poor prognosis | • Two-tailed Student’s t-test, Mann-Whitney, Spearman’s correlation test |
| Unspecified CHF | |||
| Ahmad et al.14 | • ↑TMAO#, acetate levels#, sCD14#, sCD163#, alanine#, creatinine#, glucose#, glycine#, glutamic acid#, lactic acid# • ↓3-hydroxybutyric acid#, acetic acid#, glutamine#, histidine#, LDL#, HDL#, total plasma cholesterol subfractions# • (=) propionate, butyrate, SCFAs |
• Reduction in Lachnospira, increase the plasma acetate levels in HF is compensatory response to protect the failing heart# • Increase in TMAO levels associated with congestion and impaired intestinal barrier |
• NR/Wilcoxon rank-sum test (WRST) |
| Huang et al.15 | • ↑long-chain fatty acids (cholesterone, nonadecanoic acid)#, ↓amino acid products (GABA, serotonin)#, anti-inflammatory mediators (abietic acid, quinic acid, linoleic acid and arbutin)# and neurotransmitters (catechin, caffeic acid, serotonin, tryptamine, beta-sitosterol and phenylethylamine)# • Disrupt maintenance of bile acid homeostasis and cholesterol metabolism, cysteine /methionine metabolism, leucine/isoleucine biosynthesis, ECM receptor interaction |
• Reduction in Anaerostipes, increase long-chain fatty acids and decrease amino acid products • Reduction in Parasutterella leads to impaired bile acid and lipid metabolism disorder • Increase in Azospirillum, Prevotella and Enterocloster showed enrichment of amino acid and ECM pathways |
• KEGG database, Hypergeometric tests/ Student’s t-test, Kruskal-Wallis test, Wilcoxon rank sum test, Spearman’s rank correlation |
| Zhang et al.21 | • ↑TMAO#, Choline-TMA-TMAO production pathway#, BNP#, PAGln# • Prevent intestinal corticosterone synthesis |
• Increase in TMAO-producing bacteria (Klebsiella), leads to increase risk of death and contribute to the development of adverse ventricular remodeling and HF • PAGln can act on the Beta-2-adrenergic receptors in the heart, resulting in hyperactivation of the sympathetic nervous system, which can exacerbate HF • Increase in Bacteroides can create arachidonic acid to prevent intestinal corticosterone synthesis and cut down intestinal corticosterone’s elevation owing to a high-salt diet, therefore, exerting a hypotensive effect |
• MultiQuant software/ Chi-square test, independent samples t-test, one-way ANOVA, Mann-Whitney U-test, Kruskal-Wallis test, Spearman’s rank correlation |
| Peng et al.16 | • ↑steroid hormone synthesis & TMAO • (=) acetic acid, propionate acid, butyric acid, caproic acid |
• Increase in Nocardiaceae, leads to increase in steroid hormone synthesis • Increase in Proteobacteria and Actinobacteria, leads to increase in TMAO • Firmicutes and Bacteroidota were positively associated with most of SCFAs production • The abundance of Chlamydiae was positively associated with caproic acid |
• Gas chromatography-mass spectrometry/Student’s t-test, Mann Whitney U-test, Spearman’s rank correlation |
| Wang et al.18 | • ↑ethylsalicylate#, N-formyl-L-methionine#, galacturonate#, cellobiose#, sucrose#, turanose#, alpha-lactose#, lactose#, trehalose#, isomaltose#, melibiose#, 3-amino-5-hydroxybenzoate#, 1,11-undecanedicarboxylic acid#, D-glucurono-6, 3-lactone#, adenosine#, 4-acetamidobutanoate#, homocitrulline#, N-acetylglutamate#, bilirubin#, ascorbate#, mesoporphyrin IX#, 5-methoxysalicylic acid#, vanillic acid#, 2-pyrocatechuic acid#, gentisic acid#, furosemide#, IL-6#, IL-8#, TNF-α# • ↓IL-10# |
• NR | • Internal MS2 database (BiotreeDB), ELISA/ Unpaired two-tailed Student’s t-test, Spearman’s rank correlation |
#: Significant difference; (=) no change/similar; ↑ increased; ↓ decreased; BNP: brain natriuretic peptide; CHF: chronic heart failure; ECM: extracellular matrix; GABA: gamma-aminobutyric acid; HF: heart failure; HFmrEF: heart failure with mildly reduced ejection fraction; HDL: high-density protein; HFpEF: heart failure with preserved ejection fraction; HFrEF: heart failure with reduced ejection fraction; IL-6: interleukin-6; IL-8: interleukin-8; IL-10: interleukin-10; LDL: low-density protein; NR: not reported; PAGln: phenylacetylglutamine; sCD14: soluble cluster of differentiation 14; sCD163: soluble cluster of differentiation 163; SCFAs: short-chain fatty acids; TMA: trimethylamine; TMAO: trimethylamine N-oxide; TNF-α: tumor necrosis factor-alpha.
SCFAs were frequently reduced, particularly in HFpEF and mixed HF populations, largely reflecting the depletion of SCFA-producing taxa such as Ruminococcaceae, Lachnospiraceae, and Dialister. SCFAs are key metabolites that maintain gut barrier integrity, exert anti-inflammatory effects, and regulate blood pressure through vasodilation. Their depletion therefore corresponds to impaired cardioprotective signalling and may contribute to HF progression.
Several studies also reported increased lactic acid and long-chain fatty acids, alongside reductions in amino acid-related metabolites including glutamine, histidine, serotonin, and GABA, which collectively signalled a loss of anti-inflammatory and cardioprotective activity.
At the pathway level, HF patients exhibited broad metabolic dysregulation. This included enhanced degradation of carbohydrates (e.g., trehalose, pentose phosphate and glycerol) and fatty acids, as well as altered amino acid catabolism and steroid hormone synthesis. These changes were frequently driven by opportunistic taxa such as Enterobacter, Shigella, and other Proteobacteria, mechanistically linking them to intestinal inflammation and immune activation.
Overall, the metabolomic signature of HF across phenotypes is characterized by increased TMAO and lactic acid, together with reduced SCFAs and amino acid/anti-inflammatory metabolites. Functional enrichment in fatty acid, amino acid, and steroid pathways underscores a shift toward impaired cardioprotection, heightened inflammation, and maladaptive host microbiota interactions.
Summary of Main Results
This systematic review synthesizes current evidence on gut microbiota composition and gut-derived metabolites in patients with HF. Overall, the findings demonstrate consistent alterations in both microbial diversity and functional metabolism, reflecting disrupted host-microbiota interactions in HF.
In terms of gut microbiota diversity, most studies reported reduced richness and diversity in HF patients compared with HCs, irrespective of the indices applied. This pattern was particularly evident in HFpEF and mixed HF phenotypes, whereas one study of HFrEF patients observed increased diversity, suggesting potential phenotype-specific differences. Despite this heterogeneity, the overall trend supports the notion that HF is associated with diminished microbial richness and diversity.
At the compositional level, HF patients exhibited significant shifts in microbial structure compared with HCs, as confirmed by beta diversity analyses. Across studies, reductions in Firmicutes and Bacteroidota, accompanied by increases in Proteobacteria, Actinobacteria, and Bacillota, were recurrently observed at the phylum level. Family level alterations included lower abundances of Lachnospiraceae and Oscillospiraceae and higher abundances of Enterobacteriaceae, Acidaminococcaceae, and Nocardiaceae. Genera enriched in HF included Enterobacter, Shigella, Klebsiella, Streptococcus, Lactobacillus, and Akkermansia, while key health-associated taxa such as Faecalibacterium, Roseburia, and Lachnospira were consistently depleted. At the species level, increases in pathogenic or opportunistic organisms such as Yersinia enterocolitica and Ruminococcus gnavus were reported, alongside reductions in beneficial species including Eubacterium rectale and Dorea longicatena. Collectively, these taxonomic changes reflect a dysbiotic profile characterized by loss of butyrate-producing bacteria and expansion of opportunistic taxa.
Gut microbiota derived metabolites and functional pathways also showed clear alterations in HF. TMAO was the most consistently reported metabolite, with most studies identifying elevated levels in HF patients. Increased TMAO was linked to enrichment of Escherichia and Klebsiella, and further associated with adverse ventricular remodeling, greater disease severity, and poor prognosis. In contrast, SCFAs, particularly butyrate and propionate, were frequently reduced in HF, paralleling the depletion of SCFA-producing taxa such as Lachnospiraceae, Ruminococcaceae, and Dialister. This loss corresponds to impaired gut barrier integrity, diminished anti-inflammatory signaling, and reduced cardioprotective effects.
Other notable findings included elevated lactic acid and long-chain fatty acids, together with reduced levels of amino acid-derived and anti-inflammatory metabolites such as glutamine, histidine, serotonin, and GABA. These metabolic shifts collectively indicate an imbalance toward pro-inflammatory and pro-atherogenic signalling. Functional pathway analyses further highlighted this dysregulation, with enrichment of carbohydrate degradation (e.g., trehalose, glycerol, pentose phosphate), fatty acid β-oxidation, amino acid catabolism, and steroid hormone synthesis pathways in HF patients. These changes were frequently driven by Proteobacteria and other opportunistic taxa, providing mechanistic links to intestinal inflammation, immune activation, and maladaptive host microbe interactions.
Alpha and Beta Diversity Changes in HF Patients Compared to HCs
Alpha diversity describes the microbial community within a single sample, capturing both the richness and evenness of microbial species. Richness reflects the total number of distinct taxa present, while evenness refers to how uniformly these taxa are distributed. A community is considered “richer” when it harbors a greater variety of microbial species rather than being dominated by only a few. Assessing alpha diversity is particularly important in HF research, as reduced diversity may indicate gut dysbiosis, loss of beneficial microbes, and disruption of metabolic and immune homeostasis.10 Beta diversity, on the other hand, measures the variation in microbial composition between samples or groups, providing insights into how community structure shifts across populations or environments.24 Evaluating both alpha and beta diversity is therefore essential in HF, as alterations in these measures reflect changes in gut microbiota ecosystems that may influence inflammation, metabolic pathways, and overall disease progression. Specifically, reduced alpha diversity may signal an imbalance in microbial communities, while differences in beta diversity highlight how the gut microbiota profile of HF patients diverges from that of healthy individuals.25,26
In this review, Yafarova et al.19 reported an increase in microbial richness and diversity in HFrEF patients, with significant differences observed in alpha-diversity indices (Pielou, Chao1 and PD-whole-tree) between HF patients and HCs (P < 0.001). This finding may be attributed to compensatory changes or overgrowth of opportunistic bacteria in response to the altered gut environment, including factors such as intestinal congestion or modified dietary and metabolic patterns in HF.27 In contrast, some of the included studies, reported a significant reduction in microbial richness and diversity (P < 0.05).15-18,20,22 This reduction is often linked to dysbiosis, where beneficial commensal bacteria such as Faecalibacterium and Roseburia spp. are lost and replaced by a less diverse, more pathogenic microbial community like Enterobacteriaceae or other opportunistic taxa, potentially due to reduced gut perfusion and chronic inflammation in HF patients.6 This dysbiosis may lead to reduced production of SCFAs and impaired gut barrier function, contributing to HF progression.28 Supporting this trend, a recent meta-analysis by Huang et al.,29 and a systematic review by Simadibrata et al.,30 both reported reduced richness and diversity of microbial communities, accompanied by significant alterations in beta diversity. Interestingly, Zhang et al.21 found a significant decrease in microbial richness (p < 0.05) but no significant change in diversity (p > 0.05). This may suggest that while the number of distinct microbial taxa decreased, the distribution and evenness among the remaining taxa remained relatively stable, indicating a more balanced yet narrowed microbial population.
With respect to beta diversity, eight studies consistently reported significant differences in microbial composition between HF patients and HCs (p < 0.05). This indicates that, regardless of whether diversity increased or decreased, the overall microbial structure in HF patients diverges distinctly from that of healthy individuals, reflecting disease-associated shifts in metabolic and inflammatory pathways.11 By contrast, Yang et al.20 observed no statistically significant diversity differences within the HC group, highlighting that healthy gut microbiota tends to maintain stable diversity and structural balance.
Taken together, these findings suggest that HF is characterized by heterogeneous but consistently altered microbial diversity patterns, ranging from reductions in richness and evenness to compensatory expansions of opportunistic taxa. Meanwhile, healthy individuals maintain more stable and diverse microbial communities, underscoring the role of gut dysbiosis in shaping HF pathophysiology.
Gut Microbial Taxa Associated with HF Patients Compared to HCs
The human gut microbiome is increasingly regarded as a functional “organ” with key roles in metabolism, immunity, the nervous system, and endocrine regulation.31 It hosts the largest microbial community in the body, comprising trillions of microorganisms encoding more than three million microbial genes which far exceeds the ~23,000 genes of the human genome, underlining its major contribution to human health.32,33
In healthy individuals, the microbiota is dominated by strict anaerobes, which outnumber facultative anaerobes and aerobes by up to 100-fold.34 Although more than 50 bacterial phyla have been detected in the human gut, Firmicutes and Bacteroidota together represent ~90% of the total community, while Actinobacteria, Proteobacteria, and Verrucomicrobia are present in lower proportions.35,36 The Firmicutes include SCFA-producing taxa such as Clostridium, Ruminococcus, Roseburia, and Faecalibacterium, whereas Bacteroidota are mainly represented by Bacteroides and Prevotella.34,35 Actinobacteria are enriched for Bifidobacterium, while Proteobacteria and Verrucomicrobia are maintained at relatively low levels in health. The colon, harboring the highest microbial density, is particularly enriched with butyrate producers such as Anaerostipes and Butyrivibrio, which support mucosal health and host energy metabolism.35 Despite variability at the species level, functional redundancy ensures stability of key microbial functions, including SCFA production, across individuals.34 Collectively, these features define a resilient and balanced microbial ecosystem in health.
In contrast, HF is consistently associated with dysbiosis, characterized by reduced microbial diversity, depletion of commensals, and enrichment of opportunistic taxa.10,37-39 At the phylum level, HF patients generally display decreased Firmicutes and Bacteroidota with corresponding enrichment of Proteobacteria and Actinobacteria. For example, Sun et al.17 reported that Firmicutes accounted for 72.4% of the gut microbiota in HCs but only 59.5% in CHF patients, while Proteobacteria were significantly higher in patients (21.3% vs. 6.9%). Similarly, Actinobacteria were more abundant in HF (2.7% vs. 0.9%), whereas Bacteroidota were modestly higher in controls (17.7% vs. 14.9%). These results indicate that healthy individuals preserve higher levels of commensal Firmicutes and lower levels of opportunistic Proteobacteria.
At the genus level, HCs consistently maintain greater abundance of SCFA-producing taxa. In Sun et al.17 Faecalibacterium represented 22.8% of the microbiota in controls but only 10.5% in CHF patients. Conversely, pathogenic genera such as Shigella, Klebsiella, and Enterococcus were markedly reduced in controls. Other studies reinforce this protective profile, Huang et al.15 observed higher Dialister and Anaerostipes in controls,15 Zhang et al.21 reported elevated Firmicutes, and Agathobacter, while Peng et al.16 described greater abundance of Faecalibacterium, Blautia, Prevotella, Clostridia, and Peptostreptococcaceae in controls.16 Collectively, these findings underscore the preservation of diverse SCFA producers as a hallmark of healthy gut ecology.
In HF populations, the opposite trend is observed, with consistent enrichment of pathobionts. Commonly reported taxa include Shigella, Klebsiella, Streptococcus, and Akkermansia.14,16-18,20-23 Additional enriched taxa, such as Prevotella, Bacteroides, Sutterella, and Megasphaera, suggest that HF-associated dysbiosis extends beyond the simple SCFA producer versus pathobiont imbalance.15,16,22 Importantly, disease severity appears to shape microbial shifts such as Klebsiella and Lactobacillus increase in advanced HF (NYHA IV), while Bifidobacterium declines progressively with worsening disease, potentially serving as a severity marker.15,21 At the species level, HF patients show depletion of Faecalibacterium prausnitzii, a major anti-inflammatory commensal, alongside enrichment of pro-inflammatory species such as Ruminococcus gnavus and Streptococcus spp.18 Mendelian randomization and genome-wide association studies further support causality which Bacteroides dorei emerged as the most consistently HF-associated species across large biobanks, while SCFA producers such as Butyrivibrio and Faecalibacterium prausnitzii were inversely associated with HF risk.40
Taken together, these findings highlight a reproducible dysbiotic signature in HF, marked by loss of SCFA-producing commensals and enrichment of pathobionts. In contrast, HCs maintain a balanced, SCFA rich community with lower abundance of opportunistic taxa. These taxonomic differences underpin functional consequences, including reduced SCFA generation, increased TMAO production, and altered BA metabolism, thereby providing mechanistic links between the gut microbiome and HF progression.
Alterations in Gut-Derived Metabolites and Functional Pathways in HF Patients Compared to HCs
This systematic review highlights that patients with HF consistently display alterations in gut-derived metabolites and functional pathways compared with HCs. These interconnected changes are summarized in Figure 2, which provides an overview of microbial metabolites implicated in HF. To complement this, Figure 3 illustrates the contrasting microbial and metabolic profiles in HCs, characterized by preserved eubiosis, intact gut barrier function, and balanced metabolite production. Together, Figures 2 and 3 highlight the dysbiosis-eubiosis continuum that underpins gut-heart interactions in health and disease.
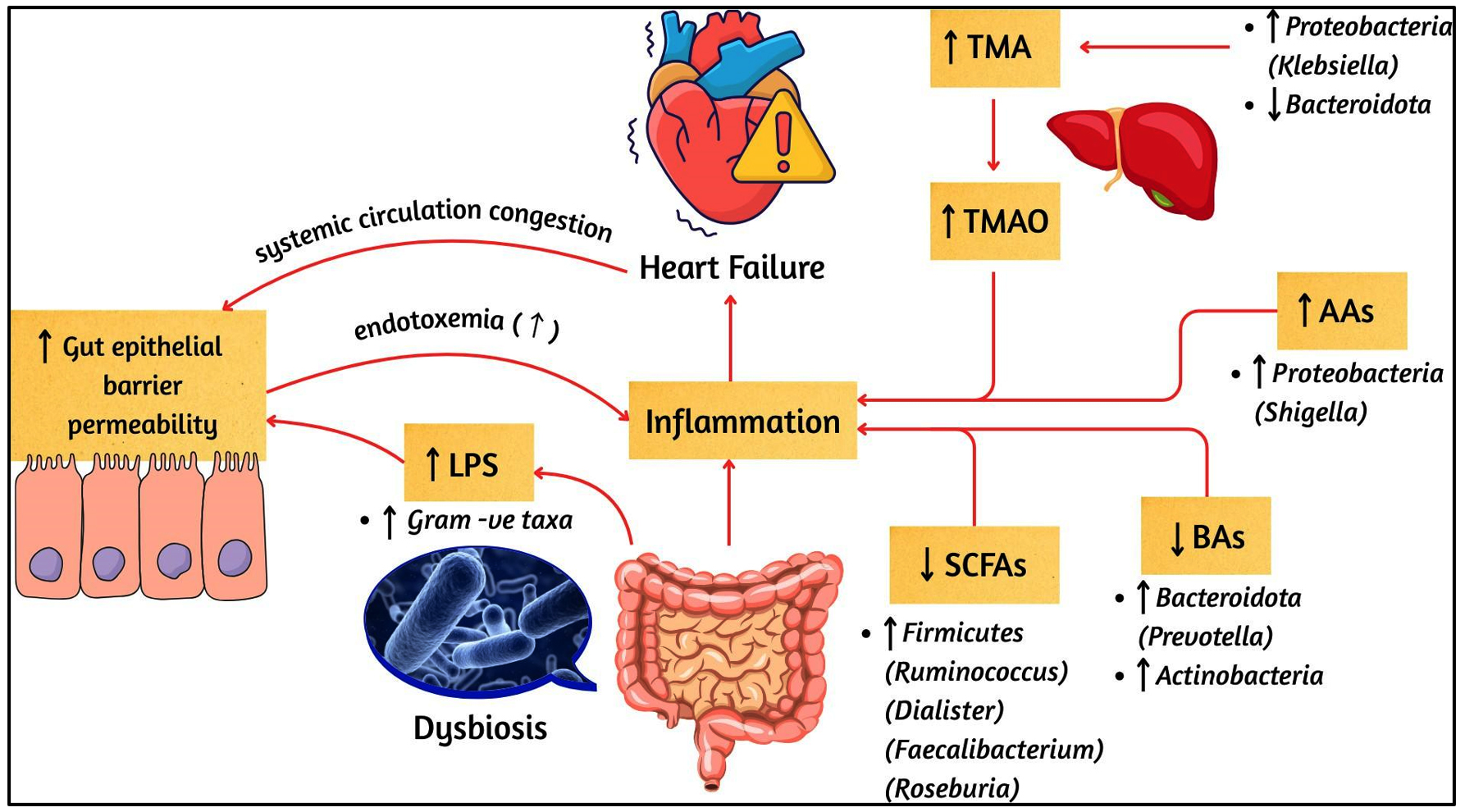
Figure 2. Gut Microbial Dysbiosis and Altered Metabolite Pathways in HF
↑: increase; ↓: decrease; AAs: amino acids; BAs: bile acids; LPS: lipopolysaccharides; SCFAs: short-chain fatty acids; TMA: trimethylamine; TMAO: trimethylamine N-oxide. Microbial genera are shown in brackets after their respective phyla
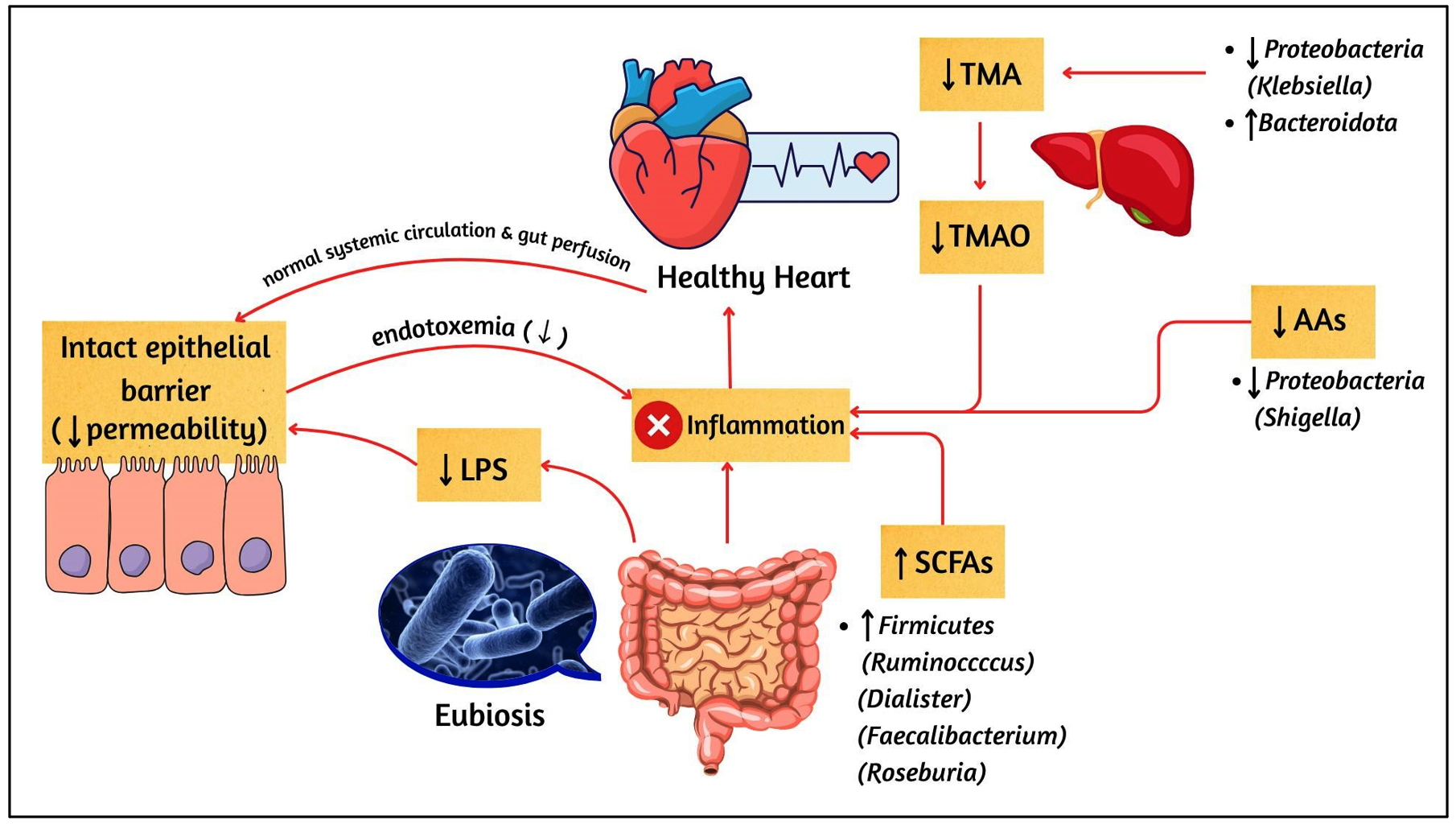
Figure 3. Gut Microbial Eubiosis and Balanced Metabolite Pathways in HCs
↑: increase; ↓: decrease; AAs: amino acids; LPS: lipopolysaccharides; SCFAs: short-chain fatty acids; TMA: trimethylamine; TMAO: trimethylamine N-oxide. Microbial genera are shown in brackets after their respective phyla
Among these, TMAO emerged as the most frequently reported metabolite, mentioned in five of the included studies, making it the central focus in the literature. TMAO is a small odourless molecule with molecular mass of 75.11 g/mol.41 TMAO is generated from dietary precursors such as choline, L-carnitine, betaine, and phosphatidylcholine which are metabolized into trimethylamine (TMA) by gut microbial enzymes such as choline-TMA lyase (cutC/D), carnitine monooxygenase (cntA/B), betaine reductase and TMAO reductase primarily produced by Firmicutes (Clostridium), Proteobacteria (Escherichia, Klebsiella) and Bacteroidota before undergo hepatic oxidation by flavin-containing monooxygenase (FMO) into TMAO.42,43 Circulating TMAO has been linked to several mechanisms relevant to HF, including vascular inflammation, oxidative stress, myocardial fibrosis, and impaired cardiac contractility.44
Most studies reported increased TMAO levels in HF patients.14,16,21,23 For example, two of the included studies21,23 associated higher TMAO with enrichment of Klebsiella and Escherichia (Proteobacteria), both known TMAO producers, and demonstrated links with more severe HF phenotypes and poor prognosis. Ahmad et al.14 further associated elevated TMAO with congestion, impaired intestinal barrier integrity, and systemic inflammation. One of the included studies, identified upregulated steroid hormone synthesis alongside increased TMAO, suggesting possible interactions with vascular tone and sodium-water balance.16 Consistent with these findings, recent meta-analysis by Li et al.,26 and systematic review by Zhang et al.,42 have reported that HF is associated with elevated circulating TMAO, supporting the concept that gut dysbiosis may contribute to maladaptive cardiovascular remodeling. In contrast, Yafarova et al.19 reported lower TMAO levels in HFrEF patients, which correlated positively with carbohydrate intake and the abundance of Firmicutes (Lachnospiraceae). The authors suggested that widespread use of statins and aspirin in that population may have lowered circulating TMAO levels. Collectively, these findings indicate that while TMAO is consistently emphasized, its levels vary with diet, medication, and clinical subtype, but overall evidence supports its role as a biomarker of gut-heart axis dysregulation.
Beyond TMAO, multiple other metabolites and pathways were dysregulated across the included studies. SCFAs, mainly acetate, propionate, and butyrate, are important products of bacterial fermentation of resistant starches and dietary fibers. They are primarily generated by Firmicutes (Faecalibacterium, Roseburia, Ruminococcus), Bacteroidota (Bacteroides), and Actinobacteria (Bifidobacterium).40,45 Two of the included studies, conducted in HFpEF and mixed HF populations, consistently reported lower SCFA levels in patients compared with controls, reflecting the loss of SCFA-producing bacteria.17,22 One of the included studies however, observed no significant changes in SCFA or butyrate levels, possibly due to small sample sizes, medication use, or the confounding effects of COVID-19 in their study population.14 Consistently, metagenomic analyses have shown reduced abundance of butyrate-producing genes in HF patients.46 In contrast, HCs as illustrated in Figure 3 exhibited preserved enrichment of SCFA producers, consistent with a protective metabolic profile.
Mechanistic studies help explain why these changes are clinically relevant. In the gut, SCFAs strengthen the intestinal barrier by enhancing tight junctions and stimulating mucus production, thereby preventing translocation of harmful microbial products such as LPS into circulation.39 Butyrate also supports colonocyte energy metabolism through PPAR-γ signaling, while butyrate and propionate promote the expansion of regulatory T cells, maintaining immune balance.47,48 Beyond the gut, SCFAs act on the cardiovascular system by activating G protein-coupled receptors (GPR41, GPR43, and GPR109A), influencing vascular tone and blood pressure, and regulating inflammatory and metabolic pathways through epigenetic mechanisms.49,50
Animal studies provide further support for these protective roles. In murine models, supplementation with SCFAs or fiber-rich diets reduced hypertension, cardiac hypertrophy, and fibrosis,51 while SCFA administration after myocardial infarction promoted favorable immune remodeling in the heart and improved recovery.11 Conversely, experimental models of SCFA depletion demonstrated gut dysbiosis, systemic inflammation, vascular dysfunction, and insulin resistance, all of which exacerbate HF progression.38,52 Together, Figure 2 shows reduced SCFA signaling in HF, whereas Figure 3 illustrates preserved SCFA production and systemic benefits in HCs.
Several studies also highlighted disruptions in amino acid metabolism. Protective metabolites such as glutamine, histidine, GABA, and serotonin were consistently reduced14,15 loss of these metabolites may reduce antioxidant capacity, impair vascular regulation, and worsen systemic inflammation. Conversely, harmful metabolites such as phenylacetylglutamine (PAGln) were elevated, with Zhang et al.21 showing that PAGln promotes sympathetic overactivation by acting on β-adrenergic receptors. Indoxyl sulfate (IS), another harmful metabolite, showed no significant change in,19 although previous studies linked IS with cardiovascular events.53 A study by Yang et al.20 expanded on these findings by identifying upregulation of amino acid catabolic pathways, including proline, lysine, phenylalanine, and arginine degradation, driven by Proteobacteria (Enterobacter, Shigella). These alterations were linked to intestinal inflammation, metabolic stress, and systemic immune activation. In contrast, HCs showed preserved amino acid metabolism, with enrichment of protective metabolites (glutamine, histidine, and serotonin), reflecting balanced microbial and host pathways.
Other pathways were also dysregulated, particularly those involving LPS and BA metabolism. Evidence shows that LPS, a structural component of Gram-negative bacterial membranes, is a central mediator of gut-heart interactions in HF. One of the included studies reported that HF patients had higher circulating levels of soluble CD14 (sCD14) and soluble CD163 (sCD163), both well-established markers of LPS exposure and monocyte/macrophage activation.14 Elevated sCD14 has previously been linked to adverse cardiomyopathy outcomes and increased mortality risk, while sCD163 reflects macrophage activation and has been shown to predict incident cardiovascular events.54,55 Together with other evidence, these findings suggest that sCD14 and sCD163 are not only markers of microbial translocation but also promising biomarkers of HF progression.56,57
Mechanistically, LPS engages toll-like receptor 4 (TLR4) on immune and endothelial cells, activating NF-κB signaling and driving release of pro-inflammatory cytokines such as TNF-α, IL-6, and IL-8, while suppressing the anti-inflammatory cytokine IL-10.58 LPS also interacts directly with CD14, amplifying these responses.59 Wang et al.18 provides direct evidence for this pathway, reporting elevated IL-6, IL-8, and TNF-α alongside reduced IL-10 in HF patients, consistent with gut-derived LPS signalling and systemic inflammation. Importantly, not all LPS molecules exert equal effects: Bacteroides-derived LPS is relatively weak, Escherichia coli LPS is highly pro-inflammatory, and Prevotella-derived LPS displays intermediate potency. Given that HF patients show microbial shifts toward Enterobacteriaceae and other Gram-negative taxa,20 the resulting exposure to more immunostimulatory LPS chemotypes may help explain the strong inflammatory profile observed in HF (Figure 2). Conversely, HCs (Figure 3) exhibited intact epithelial barrier function, reduced LPS translocation, and immune homeostasis.
BA metabolism was also disrupted, reflecting the interplay between the gut microbiome, host lipid metabolism, and cardiac function. One of the included studies, reported increased long-chain fatty acids and reduced anti-inflammatory metabolites and neurotransmitters, consistent with impaired BA and cholesterol homeostasis.15 The depletion of Parasutterella (Proteobacteria), a genus crucial for BA metabolism, was linked to disrupted cholesterol pathways, while enrichment of Prevotella (Bacteroidota) and Enterocloster (Firmicutes) suggested compensatory activation of amino acid and extracellular matrix (ECM) related metabolism. Similarly, Peng et al.16 observed that expansion of Proteobacteria, Actinobacteria, and Nocardiaceae was associated with increased steroid hormone synthesis and elevated TMAO, reinforcing the convergence of BA, lipid, and TMAO related pathways in HF.
These observations align with earlier reports showing an altered BA pool in chronic HF, characterized by reduced circulating primary BAs and elevated secondary BAs.39 Such changes are clinically important: secondary BAs modulate vascular tone, cardiac contractility, and immune responses through receptors such as FXR and TGR5, while higher secondary to primary BA ratios have been linked to systemic inflammation and worse outcomes in HF. Conversely, pharmacological modulation of BAs may offer therapeutic benefits. For example, ursodeoxycholate, a secondary BA, improved peripheral blood flow in HF patients, possibly through its endotoxin-neutralizing properties.60
Conversely, in HCs, a diverse and balanced microbial community maintains a robust intestinal epithelial barrier, which effectively prevents the translocation of inflammatory agents like LPS into the systemic circulation61 as presented in Figure 3. This barrier integrity reduces endotoxemia and systemic inflammation, which is fundamental to a healthy cardiovascular system.27 A healthy gut is characterized by low levels of opportunistic pathogens from the Proteobacteria and Bacteroidota phyla.16 This composition is associated with a decrease in the production of harmful metabolites. For instance, the low abundance of Klebsiella and Bacteroidota contributes to a reduction in the production of TMA and its liver-derived product, TMAO, which are known to be pro-atherogenic and pro-inflammatory. Additionally, the decreased abundance of Shigella is linked to a reduction in specific amino acids, further highlighting a shift away from a disease-promoting metabolic profile.17 Other than that, a healthy gut also rich in beneficial bacteria like Faecalibacterium, Roseburia, and Ruminococcus (from the Firmicutes phylum), produces SCFAs that not only nourish the colonocytes but also exert potent anti-inflammatory effects.62
HCs also showed broader and more balanced metabolic capacity. One of the included studies, reported enrichment of metabolites such as citramalate, pyroglutamate, inosine, riboflavin, and succinate in HCs, reflecting preserved amino acid, vitamin, and energy metabolism.18 Functional predictions revealed enrichment of replication/repair and terpenoid/polyketide pathways in HCs. This aligns with evidence that healthy gut microbiota produces SCFAs, vitamins (K, B12, folic acid, biotin), endocannabinoids, and BA derivatives that support metabolic and immune balance.63 Importantly, eubiosis in HCs prevents overgrowth of pathobionts through competitive exclusion, antimicrobial peptides, and SCFA production, maintaining resilience and reducing inflammation.
Collectively, these findings demonstrate that HF is not driven by a single metabolite but by a network of disrupted microbial functions. TMAO remains the most consistent biomarker, but reductions in SCFAs, accumulation of harmful metabolites such as PAGln, LPS-driven inflammation, and BA and amino acids dysregulation all contribute to impaired energy metabolism, immune imbalance, and vascular dysfunction (Figure 2). Comparisons with HCs emphasize the preservation of diverse metabolic pathways in eubiosis (Figure 3), contrasting with the restricted and pro-inflammatory metabolite profile in HF.
Key points summary
In summary, this systematic review highlights several consistent patterns linking gut microbiota dysbiosis with heart failure pathophysiology:
- Reduced microbial diversity and loss of commensal SCFA-producing taxa (Faecalibacterium, Roseburia, Lachnospira) are common in HF, contributing to impaired gut barrier integrity and inflammation.
- Enrichment of pathogenic and TMAO-producing bacteria (Klebsiella, Escherichia, Streptococcus) is associated with elevated TMAO levels, adverse cardiac remodeling, and disease progression.
- Disruption of key metabolic pathways, including decreased SCFAs and increased pro-inflammatory metabolites (e.g., PAGIn, LPS), underpins immune activation and endothelial dysfunction in HF.
- Altered bile acid and amino acid metabolism further reflects systemic metabolic dysregulation linking the gut and heart.
- Comparisons with healthy controls consistently show preserved microbial richness, balanced metabolite production, and reduced inflammation, emphasizing the gut-heart axis as a critical therapeutic target.
Collectively, these findings demonstrate that HF-related gut dysbiosis is multifactorial, involving microbial, metabolic, and immunological alterations. Future studies should employ longitudinal and multi-omics approaches to clarify causal mechanisms and evaluate microbiota-targeted interventions for HF management.
Limitations
This review has several important limitations. First, only ten studies met the eligibility criteria, representing just four countries, with more than half (six studies) conducted in China. This limited geographical diversity reduces the generalizability of the findings and makes direct comparisons across studies challenging.
Second, heterogeneity in study methods, particularly in sample collection, storage, processing, and sequencing techniques, may have influenced the reported microbial profiles and metabolites, thereby limiting comparability.
Third, adjustment for potential confounders varied considerably. While some studies implemented comprehensive adjustments14,17,20 several did not adjust for key variables such as age, sex, comorbidities, medications, or lifestyle factors.16,18,19 Others only applied partial adjustments, such as controlling for BMI, smoking, or alcohol use but omitting diet and medication data.15,21 In two studies, medication and lifestyle information were not reported at all.19,22 Because gut microbiota is highly sensitive to intrinsic factors (e.g., age, comorbidities, HF severity) and extrinsic factors (e.g., diet, medication use, cultural practices), these uncontrolled variables may have significantly biased the associations observed.
Fourth, evidence for HF subtypes was limited. Only one study examined HFrEF exclusively,19 one focused on HFpEF,22 three included mixed subtypes17,20,23 and none directly investigated HFmrEF. Consequently, pooling findings across HF subtypes may mask clinically relevant differences. Current evidence suggests that gut dysbiosis may follow distinct mechanistic pathways across HF phenotypes. In HFrEF, impaired cardiac output can lead to intestinal hypoperfusion and ischemia, increasing permeability and favoring pathogenic bacterial overgrowth, which amplifies systemic inflammation.64 In contrast, HFpEF is primarily driven by endothelial dysfunction, with gut involvement typically arising in the context of right-sided congestion.65 Dysbiosis is therefore expected to differ between HFrEF and HFpEF.66 HFmrEF remains poorly characterized, with no human studies directly exploring its gut microbiome profile. This gap in the literature limits interpretation of subtype-specific changes and underscores the need for future studies to clarify whether gut microbiome alterations contribute differently to HFrEF, HFpEF, and HFmrEF.
Fifth, the predominance of cross-sectional designs (8 of 10 studies) prevents any inference regarding temporality or causality. It remains unclear whether dysbiosis contributes to HF onset and progression, or arises as a consequence of the disease and its treatment. Only two cohort studies were available, and even these had limited follow-up or single-center designs, restricting their ability to capture longitudinal changes. Prospective, multi-center cohort studies are required to establish causal links between gut microbiota and HF outcomes.
Finally, several limitations of the review process itself should be acknowledged. The literature search was confined to three major databases (PubMed, Scopus, Web of Science), and additional studies indexed elsewhere or in the grey literature may have been missed. Restricting the review to English-language publications introduces a risk of language bias, while limiting the timeframe to 2021-2025 may have excluded earlier relevant studies. By including only peer-reviewed original research articles, we also cannot exclude the possibility of publication bias. For quality assessment, we applied the Newcastle-Ottawa Scale, which is widely used but has restricted sensitivity in capturing unique sources of bias in microbiome research. Lastly, because of the small number and heterogeneity of studies, we synthesized the data narratively rather than performing a meta-analysis, limiting our ability to quantify pooled effect sizes and formally evaluate heterogeneity.
Research gaps
Several studies have reported gut microbiota dysbiosis in HF, including reduced microbial diversity and loss of SCFA producing taxa. However, most available studies focus on taxonomic alterations without integrating functional and metabolite pathways, leading to a fragmented understanding of the gut-heart axis.66 Existing research is further limited by small sample sizes, heterogeneous methodologies45 and inadequate adjustment for key confounders such as diet, medications, and comorbidities.16,18,19 Most available studies are cross-sectional, with limited longitudinal research to show changes over time, and insufficient evidence describing phenotype-specific alterations, particularly in HFmrEF.56 Moreover, lack of systematic review has synthesized recent evidence (2021-2025) to provide an updated and comprehensive perspective. This gap limits the development of targeted microbiota-based therapies and hinders the translation of gut microbiome research into clinical HF management.
Implications
The evidence linking gut microbiome dysbiosis with HF underscores opportunities for therapeutic innovation. Depletion of SCFA producing taxa and enrichment of TMAO-producing bacteria suggest that dietary modulation, prebiotic and probiotic supplementation, and postbiotic therapies may help restore microbial balance and attenuate systemic inflammation.67 Importantly, these findings also indicate that therapeutic strategies should target restoration of specific commensal genera such as Faecalibacterium and Roseburia, while reducing overgrowth of Klebsiella and Escherichia that drive TMAO production. Beyond clinical practice, these findings emphasize the importance of integrating cardiology, microbiology, and nutrition expertise to develop microbiota targeted strategies, supported by longitudinal studies and standardized methodologies.
Building on these insights, incorporation of microbiome profiling into HF risk assessment and disease monitoring could enable early identification of dysbiosis-related phenotypes, guiding personalized nutritional or probiotic interventions. Clinically, endpoints such as circulating TMAO concentrations, fecal SCFA levels, systematic inflammatory markers (e.g., IL-6, TNF-α), and echocardiographic measures of ventricular function should be considered in interventional trials to ensure measurable translational outcomes. Integration of gut microbial biomarkers alongside conventional cardiac indicators (NT-proBNP, troponins) could improve diagnostic precision and risk stratification in clinical cardiology. Furthermore, translation of microbiota-targeted therapies into clinical use will require harmonized regulatory and safety frameworks, particularly for high-impact interventions such as fecal microbiota transplantation.
At a broader level, health policies promoting fiber-rich, minimally processed diets may contribute to both prevention and management of HF through favorable effects on microbial ecology, while also providing measurable targets for population-based monitoring of cardiometabolic outcomes. Incorporating microbiome considerations into public health nutrition strategies could thus represent a sustainable approach to mitigating HF burden globally.
Recommendations
Future research should prioritize well-designed, large-scale, and longitudinal studies that address the current limitations in sample size, geographical diversity, and methodological heterogeneity observed in existing evidence on gut microbiota dysbiosis in HF.68 Multi-center investigations incorporating participants from diverse ethnic and dietary backgrounds are essential to enhance the generalizability of findings.69 Standardized protocols for sample collection, processing, and sequencing should be adopted to minimize technical variability and allow for robust cross-study comparisons.70
In addition, mechanistic studies employing gnotobiotic or humanized animal models are needed to establish causal relationships between SCFA depletion, TMAO elevation, and HF progression. Identification of microbial gene clusters involving TMA/TMAO synthesis could facilitate the development of targeted inhibitors or CRISPR- based modulation strategies. Integrative multi-omics approaches, including metagenomics, metabolomics, and transcriptomics, are recommended to elucidate causal links between microbial composition, functional pathways, and HF progression, particularly focusing on key metabolites such as TMAO and SCFAs.71
In addition to compositional profiling, clinical trials should incorporate primary endpoints including reduction in plasma TMAO levels, restoration of SCFA-producing taxa abundance (Faecalibacterium, Roseburia, Bifidobacterium), improvements in systemic inflammation (e.g., reduction in IL-6, CRP), and cardiac outcomes such as left ventricular ejection fraction, NT-proBNP, and hospitalization rates. Subtype-specific analyses, especially in HFmrEF, remain a critical gap and should be addressed to better define phenotype-dependent microbial signatures.72
Interventional studies exploring dietary modification, prebiotics, probiotics, synbiotics, or fecal microbiota transplantation should be rigorously designed to determine their therapeutic potential in restoring SCFA producing taxa (e.g., Faecalibacterium, Roseburia) and mitigating harmful microbial metabolites linked to adverse cardiovascular outcomes.73 Furthermore, future studies should account for key confounding factors, including medication use, comorbidities, and lifestyle, to ensure reliable associations between gut dysbiosis and HF.74 A more holistic investigation of the gut-heart axis, integrating environmental, metabolic, and immune determinants, will provide a foundation for the development of precise microbiota-targeted interventions in HF management.
This systematic review demonstrated consistent alterations in the gut microbiome of HF patients compared to HCs, marked by reduced microbial diversity, loss of SCFAs bacteria, and enrichment of pathogenic taxa linked to TMAO production. These compositional and functional shifts contribute to systemic inflammation, metabolic dysregulation, and adverse cardiac remodeling, underscoring the gut-heart axis as an important factor in HF progression. Differences across HF phenotypes suggest that gut dysbiosis may not be uniform, although evidence for HFmrEF remains lacking.
However, despite these consistent associations, the current body of evidence is insufficient to determine whether gut dysbiosis is a cause, a consequence, or a bidirectional component of heart failure. Most included studies were cross-sectional in design, limiting causal inference as they capture only a single time point without establishing temporal relationships. To clarify directionality, longitudinal cohort studies tracking microbiota changes from pre-clinical to advanced HF stages, as well as interventional trials that actively modulate gut microbiota composition (e.g., via probiotics, dietary fiber supplementation, or fecal microbiota transplantation), are warranted. Such study designs would help determine whether restoring eubiosis can prevent HF onset or progression, or merely reflect improved cardiac function following disease management. These findings emphasize the gut-heart axis as a key contributor to HF pathophysiology and highlight the urgent need for mechanistic and longitudinal studies to establish causal links and inform the development of microbiota-targeted strategies in HF management.
Additional file: Table S1-S6.
ACKNOWLEDGMENTS
The authors would like to express their sincere appreciation to Universiti Teknologi MARA (UiTM) for providing the facilities and support essential for this systematic review.
CONFLICT OF INTEREST
The authors declare that there is no conflict of interest.
AUTHORS’ CONTRIBUTION
NAMA, NNHMN, and AMM conceptualized the study. NAMA and NNHMN performed data collection, analyzed, interpretation and drafted the manuscript. AMM, JH, and NKP revised the manuscript. SHSAK, AMM and JH supervised the study. All authors read and approved the final manuscript for publication.
FUNDING
This study was supported by the Higher Institution Centre of Excellence (HICoE) research grant 600-RMC/MOHE HICoE CARE-I 5/3 (01/2025) awarded to the Cardiovascular Advancement and Research Excellence Institute (CARE Institute).
DATA AVAILABILITY
All datasets generated or analyzed during this study are included in the manuscript and/or in the supplementary files.
ETHICS STATEMENT
This article does not contain any studies on human participants or animals performed by any of the authors.
- Savarese G, Becher PM, Lund LH, Seferovic P, Rosano GMC, Coats AJS. Global burden of heart failure: a comprehensive and updated review of epidemiology. Cardiovasc Res. 2023;118(17):3272-3287.
Crossref - Hessel FP. Overview of the socio-economic consequences of heart failure. Cardiovasc Diagn Ther. 2021;11(1):254-262.
Crossref - Bozkurt B, Coats AJ, Tsutsui H, et al. Universal Definition and Classification of Heart Failure. J Card Fail. 2021;27(4):387-413.
Crossref - Palaparthi EC, Padala T, Singamaneni R, et al. Emerging Therapeutic Strategies for Heart Failure: A Comprehensive Review of Novel Pharmacological and Molecular Targets. Cureus. 2025;17(4):e81573.
Crossref - Ogunrinola GA, Oyewale JO, Oshamika OO, Olasehinde GI. The Human Microbiome and Its Impacts on Health. Int J Microbiol. 2020;2020:1-7.
Crossref - Shen Y, Fan N, Ma S, Cheng X, Yang X, Wang G. Gut Microbiota Dysbiosis: Pathogenesis, Diseases, Prevention, and Therapy. MedComm (Beijing). 2025;6(5):70168.
Crossref - Kondapalli N, Katari V, Dalal KK, Paruchuri S, Thodeti CK. Microbiota in Gut Heart Axis: Metabolites and Mechanisms in Cardiovascular Disease. Compr Physiol. 2025;15(3).
Crossref - Nesci A, Carnuccio C, Ruggieri V, et al. Gut Microbiota and Cardiovascular Disease: Evidence on the Metabolic and Inflammatory Background of a Complex Relationship. Int J Mol Sci. 2023;24(10):9087.
Crossref - Matacchione G, Piacenza F, Pimpini L, Rosati Y, Marcozzi S. The role of the gut microbiota in the onset and progression of heart failure: insights into epigenetic mechanisms and aging. Clin Epigenetics. 2024;16(1):175.
Crossref - Lupu VV, Raileanu AA, Mihai CM, et al. The Implication of the Gut Microbiome in Heart Failure. Cells. 2023;12(8):1158.
Crossref - Tang WHW, Li DY, Hazen SL. Dietary metabolism, the gut microbiome, and heart failure. Nat Rev Cardiol. 2019;16(3):137-154.
Crossref - Filardo S, Di Pietro M, Sessa R. Current progresses and challenges for microbiome research in human health: a perspective. Front Cell Infect Microbiol. 2024;14:1377012.
Crossref - Sorboni SG, Moghaddam HS, Jafarzadeh-Esfehani R, Soleimanpour S. A Comprehensive Review on the Role of the Gut Microbiome in Human Neurological Disorders. Clin Microbiol Rev. 2022;35(1):e0033820.
Crossref - Ahmad AF, Caparros-Martin JA, Gray N, et al. Insights into the associations between the gut microbiome, its metabolites, and heart failure. Am J Physiol Heart Circ Physiol. 2023;325(6):H1325-H1336.
Crossref - Huang K, Duan J, Wang R, et al. Landscape of gut microbiota and metabolites and their interaction in comorbid heart failure and depressive symptoms: a random forest analysis study. mSystems. 2023;8(6):e0051523.
Crossref - Peng J, Gong H, Lyu X, et al. Characteristics of the fecal microbiome and metabolome in older patients with heart failure and sarcopenia. Front Cell Infect Microbiol. 2023;13:1127041.
Crossref - Sun W, Du D, Fu T, Han Y, Li P, Ju H. Alterations of the Gut Microbiota in Patients With Severe Chronic Heart Failure. Front Microbiol. 2022;12:813289.
Crossref - Wang Z, Cai Z, Ferrari MW, et al. The Correlation between Gut Microbiota and Serum Metabolomic in Elderly Patients with Chronic Heart Failure. Mediators Inflamm. 2021;2021(1):5587428.
Crossref - Yafarova AA, Dementeva EV, Zlobovskaya OA, et al. Gut Microbiota and Metabolic Alterations Associated with Heart Failure and Coronary Artery Disease. Int J Mol Sci. 2024;25(20):11295.
Crossref - Yang C, Li X, Hu M, Li T, Jiang L, Zhang Y. Gut Microbiota as Predictive Biomarker for Chronic Heart Failure in Patients with Different Nutritional Risk. J Cardiovasc Transl Res. 2024;17(6):1240-1257.
Crossref - Zhang Z, Cai B, Sun Y, Deng H, Wang H, Qiao Z. Alteration of the gut microbiota and metabolite phenylacetylglutamine in patients with severe chronic heart failure. Front Cardiovasc Med. 2023;9:1076806.
Crossref - Beale AL, O’Donnell JA, Nakai ME, et al. The Gut Microbiome of Heart Failure With Preserved Ejection Fraction. J Am Heart Assoc. 2021;10(13):e020654.
Crossref - Emoto T, Hayashi T, Tabata T, et al. Metagenomic analysis of gut microbiota reveals its role in trimethylamine metabolism in heart failure. Int J Cardiol. 2021;338:138-142.
Crossref - Cassol I, Ibaסez M, Bustamante JP. Key features and guidelines for the application of microbial alpha diversity metrics. Sci Rep. 2025;15(1):622.
Crossref - Cienkowski K, Cienkowska A, Kupczynska K, Bielecka-Dabrowa A. The Role of Gut Microbiota and Its Metabolites in Patients with Heart Failure. Biomedicines. 2024;12(4):894.
Crossref - Li X, Fan Z, Cui J, et al. Trimethylamine N-Oxide in Heart Failure: A Meta-Analysis of Prognostic Value. Front Cardiovasc Med. 2022;9:817396.
Crossref - Acevedo-Roman A, Pagan-Zayas N, Velazquez-Rivera LI, Torres-Ventura AC, Godoy-Vitorino F. Insights into Gut Dysbiosis: Inflammatory Diseases, Obesity, and Restoration Approaches. Int J Mol Sci. 2024;25(17):9715.
Crossref - Chulenbayeva L, Issilbayeva A, Sailybayeva A, Bekbossynova M, Kozhakhmetov S, Kushugulova A. Short-Chain Fatty Acids and Their Metabolic Interactions in Heart Failure. Biomedicines. 2025;13(2):343.
Crossref - Huang J, Lin Y, Ding X, et al. Alteration of the gut microbiome in patients with heart failure: A systematic review and meta-analysis. Microb Pathog. 2024;192:106647.
Crossref - Simadibrata DM, Auliani S, Widyastuti PA, et al. The Gut Microbiota Profile in Heart Failure Patients: A Systematic Review. J Gastrointestin Liver Dis. 2023;32(3):393-401.
Crossref - Bozomitu L, Miron I, Adam Raileanu A, et al. The Gut Microbiome and Its Implication in the Mucosal Digestive Disorders. Biomedicines. 2022;10(12):3117.
Crossref - Di Tommaso N, Gasbarrini A, Ponziani FR. Intestinal Barrier in Human Health and Disease. Int J Environ Res Public Health. 2021;18(23):12836.
Crossref - Dicks LMT. Gut Bacteria Provide Genetic and Molecular Reporter Systems to Identify Specific Diseases. Int J Mol Sci. 2024;25(8):4431.
Crossref - Bull MJ, Plummer NT. Part 1: The Human Gut Microbiome in Health and Disease. Integr Med (Encinitas). 2014;13(6):17-22.
- Portincasa P, Khalil M, Graziani A, et al. Gut microbes in metabolic disturbances. Promising role for therapeutic manipulations? Eur J Intern Med. 2024;119:13-30.
Crossref - Rinninella E, Raoul P, Cintoni M, et al. What is the Healthy Gut Microbiota Composition? A Changing Ecosystem across Age, Environment, Diet, and Diseases. Microorganisms. 2019;7(1):14.
Crossref - Kummen M, Mayerhofer CCK, Vestad B, et al. Gut Microbiota Signature in Heart Failure Defined From Profiling of 2 Independent Cohorts. J Am Coll Cardiol. 2018;71(10):1184-1186.
Crossref - Cui X, Ye L, Li J, et al. Metagenomic and metabolomic analyses unveil dysbiosis of gut microbiota in chronic heart failure patients. Sci Rep. 2018;8(1):635.
Crossref - Mayerhofer CCK, Kummen M, Holm K, et al. Low fibre intake is associated with gut microbiota alterations in chronic heart failure. ESC Heart Fail. 2020;7(2):456-466.
Crossref - Dai H, Hou T, Wang Q, et al. Causal relationships between the gut microbiome, blood lipids, and heart failure: a Mendelian randomization analysis. Eur J Prev Cardiol. 2023;30(12):1274-1282.
Crossref - Papandreou C, More M, Bellamine A. Trimethylamine N-Oxide in Relation to Cardiometabolic Health—Cause or Effect? Nutrients. 2020;12(5):1330.
Crossref - Zhang Y, Wang Y, Ke B, Du J. TMAO: how gut microbiota contributes to heart failure. Transl Res. 2021;228:109-125.
Crossref - Zhou Y, Zhang Y, Jin S, Lv J, Li M, Feng N. The gut microbiota derived metabolite trimethylamine N-oxide: Its important role in cancer and other diseases. Biomed Pharmacother. 2024;177:117031.
Crossref - Caradonna E, Abate F, Schiano E, et al. Trimethylamine-N-Oxide (TMAO) as a Rising-Star Metabolite: Implications for Human Health. Metabolites. 2025;15(4):220.
Crossref - Epelde F. Heterogeneity in Heart Failure with Preserved Ejection Fraction: A Systematic Review of Phenotypic Classifications and Clinical Implications. J Clin Med. 2025;14(14):4820.
Crossref - Wu J, Qiu M, Sun L, et al. a-Linolenic Acid and Risk of Heart Failure: A Meta-Analysis. Front Cardiovasc Med. 2022;8.
Crossref - Byndloss MX, Olsan EE, Rivera-Chavez F, et al. Microbiota-activated PPAR-g signaling inhibits dysbiotic Enterobacteriaceae expansion. Science. 2017;357(6351):570-575.
Crossref - Bartolomaeus H, Balogh A, Yakoub M, et al. Short-Chain Fatty Acid Propionate Protects From Hypertensive Cardiovascular Damage. Circulation. 2019;139(11):1407-1421.
Crossref - Marques FZ, Nelson E, Chu PY, et al. High-Fiber Diet and Acetate Supplementation Change the Gut Microbiota and Prevent the Development of Hypertension and Heart Failure in Hypertensive Mice. Circulation. 2017;135(10):964-977.
Crossref - Pluznick J. A novel SCFA receptor, the microbiota, and blood pressure regulation. Gut Microbes. 2014;5(2):202-207.
Crossref - Kaye DM, Shihata WA, Jama HA, et al. Deficiency of Prebiotic Fiber and Insufficient Signaling Through Gut Metabolite-Sensing Receptors Leads to Cardiovascular Disease. Circulation. 2020;141(17):1393-1403.
Crossref - Luedde M, Winkler T, Heinsen F, et al. Heart failure is associated with depletion of core intestinal microbiota. ESC Heart Fail. 2017;4(3):282-290.
Crossref - Imazu M, Takahama H, Shindo K, et al. A Pathophysiological Role of Plasma Indoxyl Sulfate in Patients with Heart Failure. Int J Gerontol. 2017;11(2):62-66.
Crossref - Verbree Willemsen L, Zhang Y, Ibrahim I, et al. Extracellular vesicle Cystatin C and CD14 are associated with both renal dysfunction and heart failure. ESC Heart Fail. 2020;7(5):2240-2249.
Crossref - Durda P, Raffield LM, Lange EM, et al. Circulating Soluble CD163, Associations With Cardiovascular Outcomes and Mortality, and Identification of Genetic Variants in Older Individuals: The Cardiovascular Health Study. J Am Heart Assoc. 2022;11(21):e024374.
Crossref - Ma C, Luo H, Fan L, Liu X, Gao C. Heart failure with preserved ejection fraction: an update on pathophysiology, diagnosis, treatment, and prognosis. Braz J Med Biol Res. 2020;53(7):9646.
Crossref - Yuzefpolskaya M, Bohn B, Nasiri M, et al. Gut microbiota, endotoxemia, inflammation, and oxidative stress in patients with heart failure, left ventricular assist device, and transplant. J Heart Lung Transplant. 2020;39(9):880-890.
Crossref - Kiers D, Leijte GP, Gerretsen J, Zwaag J, Kox M, Pickkers P. Comparison of different lots of endotoxin and evaluation of in vivo potency over time in the experimental human endotoxemia model. Innate Immun. 2019;25(1):34-45.
Crossref - Guo S, Nighot M, Al-Sadi R, Alhmoud T, Nighot P, Ma TY. Lipopolysaccharide Regulation of Intestinal Tight Junction Permeability Is Mediated by TLR4 Signal Transduction Pathway Activation of FAK and MyD88. J Immunol. 2015;195(10):4999-5010.
Crossref - von Haehling S, Schefold JC, Jankowska EA, et al. Ursodeoxycholic Acid in Patients With Chronic Heart Failure. J Am Coll Cardiol. 2012;59(6):585-592.
Crossref - Ghosh SS, Wang J, Yannie PJ, Ghosh S. Intestinal Barrier Dysfunction, LPS Translocation, and Disease Development. J Endocr Soc. 2020;4(2):bvz039.
Crossref - Facchin S, Bertin L, Bonazzi E, et al. Short-Chain Fatty Acids and Human Health: From Metabolic Pathways to Current Therapeutic Implications. Life. 2024;14(5):559.
Crossref - Di Ciaula A, Bonfrate L, Khalil M, Garruti G, Portincasa P. Contribution of the microbiome for better phenotyping of people living with obesity. Rev Endocr Metab Disord. 2023;24(5):839-870.
Crossref - Ferro F, Spelat R, Valente C, Contessotto P. Understanding How Heart Metabolic Derangement Shows Differential Stage Specificity for Heart Failure with Preserved and Reduced Ejection Fraction. Biomolecules. 2022;12(7):969.
Crossref - Patel RB, Shah SJ. Drug Targets for Heart Failure with Preserved Ejection Fraction: A Mechanistic Approach and Review of Contemporary Clinical Trials. Annu Rev Pharmacol. 2018;59(1):41-63.
Crossref - Drapkina OM, Ashniev GA, Zlobovskaya OA, et al. Diversities in the Gut Microbial Patterns in Patients with Atherosclerotic Cardiovascular Diseases and Certain Heart Failure Phenotypes. Biomedicines. 2022;10(11):2762.
Crossref - Tang WHW, Kitai T, Hazen SL. Gut Microbiota in Cardiovascular Health and Disease. Circ Res. 2017;120(7):1183-1196.
Crossref - Yang SY, Han SM, Lee JY, Kim KS, Lee JE, Lee DW. Advancing Gut Microbiome Research: The Shift from Metagenomics to Multi-Omics and Future Perspectives. J Microbiol Biotechnol. 2025;35:e2412001.
Crossref - Shea L, Pesa J, Geonnotti G, Powell V, Kahn C, Peters W. Improving diversity in study participation: Patient perspectives on barriers, racial differences and the role of communities. Health Expectations. 2022;25(4):1979-1987.
Crossref - Lyte JM, Seyoum MM, Ayala D, et al. Do we need a standardized 16S rRNA gene amplicon sequencing analysis protocol for poultry microbiota research? Poult Sci. 2025;104(7):105242.
Crossref - Sabih Ur Rehman S, Nasar MI, Mesquita CS, et al. Integrative systems biology approaches for analyzing microbiome dysbiosis and species interactions. Brief Bioinform. 2025;26(4):bbaf323.
Crossref - Rosano GMC, Vitale C, Spoletini I. Precision Cardiology: Phenotype-targeted Therapies for HFmrEF and HFpEF. Int J Heart Fail. 2024;6(2):47.
Crossref - Wang Y, Yan H, Zheng Q, Sun X. Crucial functions of gut microbiota on gut–liver repair. hLife. 2025;3(8):364-385.
Crossref - Matute SP, Iyavoo S. Exploring the gut microbiota: lifestyle choices, disease associations, and personal genomics. Front Nutr. 2023;10:1225120.
Crossref
© The Author(s) 2026. Open Access. This article is distributed under the terms of the Creative Commons Attribution 4.0 International License which permits unrestricted use, sharing, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.