


ISSN: 0973-7510
E-ISSN: 2581-690X
Ethylenediaminetetraacetic acid, commonly used in micronutrient fertilizers as a chelating agent, enhances the availability of micronutrients to plants. However, its accumulation in the environment has numerous negative effects. The search for alternative chelating agents without these drawbacks is highly relevant. This study evaluated the impact of a novel micronutrient fertilizer based on the chelating agent 2-((1,2-dicarboxyethyl)amino)pentanedioic acid on the endophytic microbial community of lettuce grown in soil and hydroponic systems. The OTU count in soil was 1595, compared to 449 in hydroponics. In hydroponic systems, the application of 2-((1,2-dicarboxyethyl)amino)pentanedioic acid altered the abundance of several dominant bacterial taxa. In soil experiments, the effect of 2-((1,2-dicarboxyethyl)amino)pentanedioic acid on bacterial taxa was less pronounced. For endophytic bacterial communities, 12 bacterial OTUs were identified as core for lettuce leaves and roots regardless of the substrate. The OTU count in control soil root samples was 296, compared to 178 in hydroponic roots. The application of 2-((1,2-dicarboxyethyl)amino)pentanedioic acid led to slight shifts in the abundance of certain endophytic bacteria, especially in hydroponic systems. Non-metric multidimensional scaling analysis demonstrated that substrate type has the most significant influence on the composition of endophytic and substrate bacterial communities. The application of 2-((1,2-dicarboxyethyl)amino)pentanedioic acid did not significantly alter the composition of the communities. The results suggest that while 2-((1,2-dicarboxyethyl)amino)pentanedioic acid chelated fertilizer can influence the abundance of specific microbial taxa, it does not drastically disrupt the endophytic bacterial communities’ structure in lettuce grown in soil or hydroponics.
2-((1,2-dicarboxyethyl)amino)pentanedioic Acid, Micronutrient Fertilizers, Chelates, Ethylenediaminetetraacetic Acid, Plant Endophytic Microbiome, Bacterial Community, Soil, Hydroponics
Achieving high yields of agricultural crops necessitates comprehensive agronomic practices, including the use of various organic and mineral fertilizers. These fertilizers enhance the soil’s overall condition and provide essential nutrients to plants.1 Despite the evident benefits of fertilizers, there are significant drawbacks. Excessive and uncontrolled application of fertilizers can degrade soil structure, reduce fertility, contaminate groundwater and surface water, and increase greenhouse gas emissions from the soil.2
To enhance the efficiency of nutrient delivery to plants, synthetic micronutrient fertilizers based on chelating agents have been developed. Chelating fertilizers offer high protection against oxidation, precipitation, and immobilization. Chelation significantly improves nutrient uptake by plants, accelerates growth, strengthens plant immunity, and increases yields. Ethylenediaminetetraacetic acid (EDTA) is one of the most popular and effective chelating agents for producing micronutrient fertilizers.3 However, EDTA’s low biodegradability results in its accumulation in soils and groundwater.4 In soil, EDTA converts heavy metals into a soluble form, leading to their accumulation in plants and contamination of drinking water. High concentrations of EDTA can reduce seed germination and plant biomass, slow growth, and delay the average emergence time of seedlings. In the presence of EDTA, plants may produce fewer photosynthetic pigments and generate more reactive oxygen species, resulting in electrolyte leakage. EDTA is also known to adversely affect soil microbial communities, altering both the number and diversity of microorganisms.5
Given the aforementioned disadvantages of EDTA-based fertilizers, the search for new biodegradable chelating agents that enhance environmental safety is imperative. Alternatives like iminodisuccinic acid (IDS), EDDHA, and N,N’-bis(2-hydroxyphenyl)ethylenediamine-N,N’-diacetic acid (HBED), which is an isomer of EDDHA, are commercially available and environmentally friendly.6 In our previous work, the prospects of IGSA as such a chelate were demonstrated. The yield of lettuce treated with IGSA-based fertilizer was 1.4-1.6 times higher than with an EDTA-based fertilizer, while the biodegradability of IGSA-based fertilizer was 59% higher.6
To recommend IGSA as a full-fledged alternative to EDTA, information on the effect of IGSA chelates on plant-associated microorganisms is also needed. These microorganisms support plant function and protection and can directly influence human health, particularly when plants are consumed raw. The endophytic community of plants includes microorganisms residing in various plant parts, occupying intercellular or intracellular spaces.6 The composition of a plant’s microbial community is derived from seed microorganisms and those recruited from the environment. Plant seed endophytes are generally highly conserved, whereas the plant’s endophytic microbial community can change during growth and development. It is also influenced by various abiotic and biotic factors, such as soil conditions, biogeography, plant genotype, and interactions among microorganisms and plants.7
The literature documents various negative effects of different fertilizers on the endophytic microbial community. For instance, nitrogen fertilizer application altered the microbial community structure in seeds, roots, leaves, and stems, affecting the relative abundance of several beneficial microorganisms, as well as affected genes involved in the nitrogen cycle in the roots.8,9 Alterations in the composition and structure of the plant endophytic microbiome can negatively impact plant immunity and resistance to infections, ultimately leading to crop losses.10
In this study, we investigated the potential of using a new chelating agent, IGSA, to develop a plant micronutrient fertilizer. The objective was to evaluate the effect of IGSA-based fertilizer on endophytic microorganisms of lettuce, cultivated on different substrates soil and hydroponics.
Experimental design
The IGSA was synthesized at the Chemical Institute of Kazan Federal University (Russia). The structural formula and characteristics of IGSA, as well as the IGSA-based microfertilizer, are described in our previous work.6 Lettuce plants (Lactuca sativa L.) were grown in two substrate systems-hydroponics and soil in a greenhouse. The preparation of seeds, seedlings, and the design of soil and hydroponics experiments are detailed in.6 In the hydroponics experiment, IGSA-based fertilizer was introduced into the water. In the soil experiment, it was applied in two ways: foliar (on leaf surfaces) and root application (at the plant roots). Both experiments (soil and hydroponics) included control variants without the application of IGSA-based fertilizer (Figure 1).
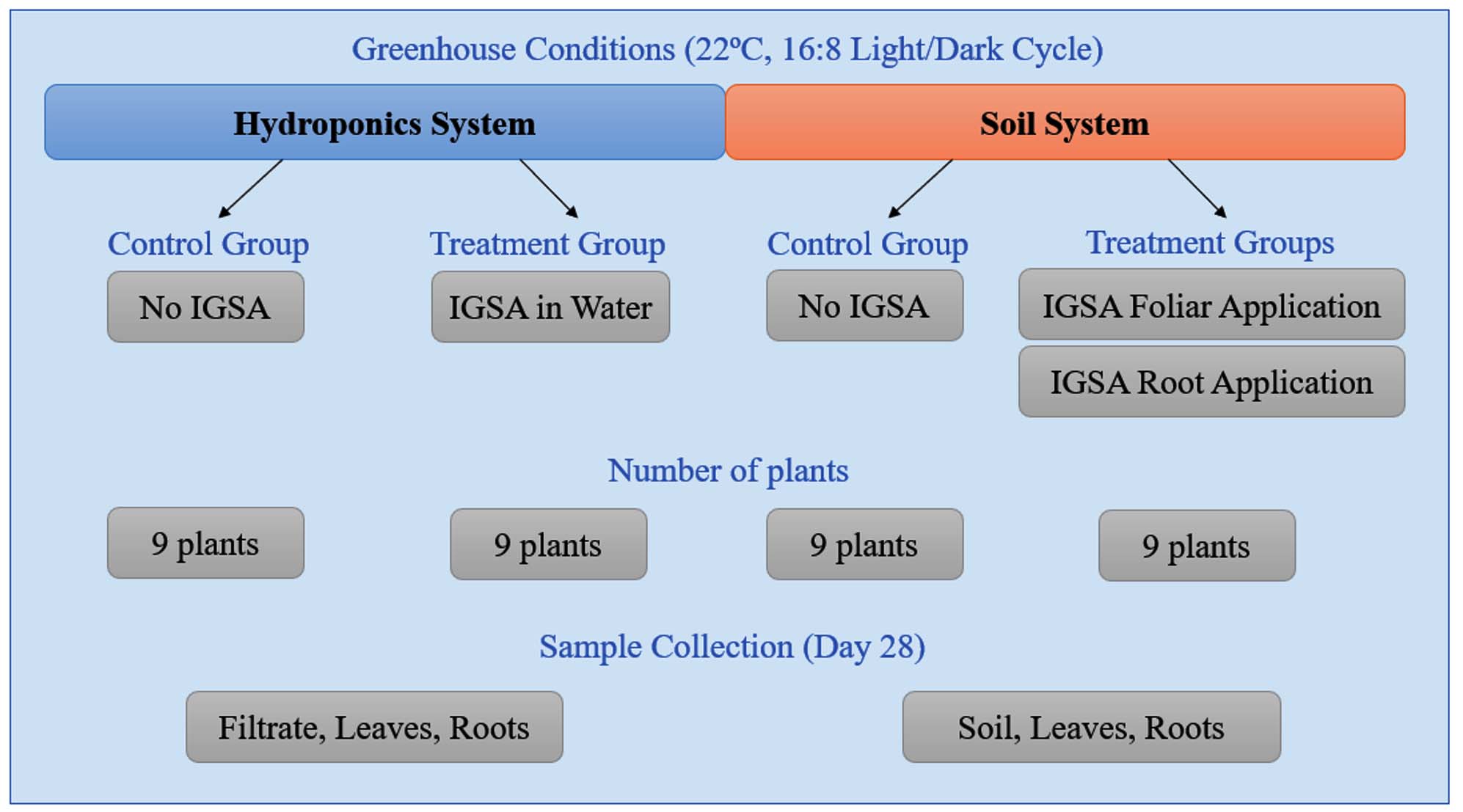
Figure 1. Scheme of the hydroponic and soil experiment
The characteristics of the soil, the content of micronutrients in the hydroponic solution, and the concentration of IGSA-based microfertilizer applied are described in Brusko et al.6 Plants were cultivated in a greenhouse at 22 °C with a 16:8 hour light/dark cycle for 28 days. On the 28th day of vegetation, plants were harvested from the substrate. Filtrate, soil, leaves, and roots from each container were immediately used for bacterial DNA extraction.
DNA extraction and sequencing
DNA extraction from plant and soil samples was performed using the FastDNA Spin Kit for Soil (MP Bio., Irvine, CA, USA) following the manufacturer’s instructions. DNA purification was conducted using the QIAquick PCR Purification Kit (Qiagen, Dusseldorf, Germany).
Sequencing was conducted according to the established protocol (https://support.illumina.com/documents/documentation/chemistry_documentation/16s/16s-metagenomic-library-prep-guide-15044223-b.pdf). Sequencing of the bacterial community was carried out on an Illumina platform (Illumina, San Diego, CA, USA). 16S rRNA sequencing data were analyzed using the Quantitative Insights Into Microbial Ecology (QIIME, East Lansing, Michigan, USA) platform, version 1.6.0.11 Representative sequences were aligned using the PyNAST algorithm.12 Within the high-throughput sequencing data analysis it was observed 1,777,897 qualified sequences from all samples for the bacterial microbial community. The total number of chimeras excluded from the analysis was 64,990 (Supplementary Table S1).
Statistical analysis
All measurements were conducted in at least three replicates. Each container for plant cultivation housed nine plants. Statistical analyses of the results were performed using the R Statistical Software package (version 3.6.1) and Microsoft Office Excel 2010 (Redmond, WA, USA) (R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, 2019). All graphical data are presented as mean values with standard deviations. The Mann–Whitney test was employed to determine the significance of differences at α = 0.05. The relative similarity of bacterial communities was assessed using non-metric multidimensional scaling (NMDS) based on the Bray-Curtis coefficient.13 Shannon and Simpson indices were calculated to measure the alpha diversity of microbial communities.14,15 Venn diagrams illustrating shared and unique bacterial OTUs of microbial community samples were generated using an online tool (http://bioinformatics.psb.ugent.be/webtools/Venn/).
The use of various substances in plant cultivation, targeting different purposes such as nutrition, disease protection, pest control, etc., is widespread in agriculture worldwide.16,17 However, alongside the positive impact on the growth and development of crops, such substances can also have negative effects, particularly on the microbial communities in both soil and plant endophytes.18,19 Given that IGSA, a chelated fertilizer, is a new product and there is insufficient data on its effects on microbial communities, this study was conducted.
Influence of IGSA on bacterial communities of substrates – hydroponics and soil
In the first stage, we assessed whether the application of IGSA affected the number of operational taxonomic units (OTUs) and their diversity in the substrate and the endosphere of lettuce. In hydroponic cultivation of lettuce, the control samples (without IGSA) had OTU counts in filtrate, leaves, and roots of 449, 51, and 178, respectively. The Simpson and Shannon indices, characterizing alpha diversity of microbial communities in the control filtrate samples, were 0.96 and 4.03, in leaves – 0.95 and 3.53, and in roots – 0.9 and 3.1. Figure 2 shows the OTU counts and the Simpson and Shannon indices expressed as percentages of the control. The application of IGSA did not significantly affect these parameters.
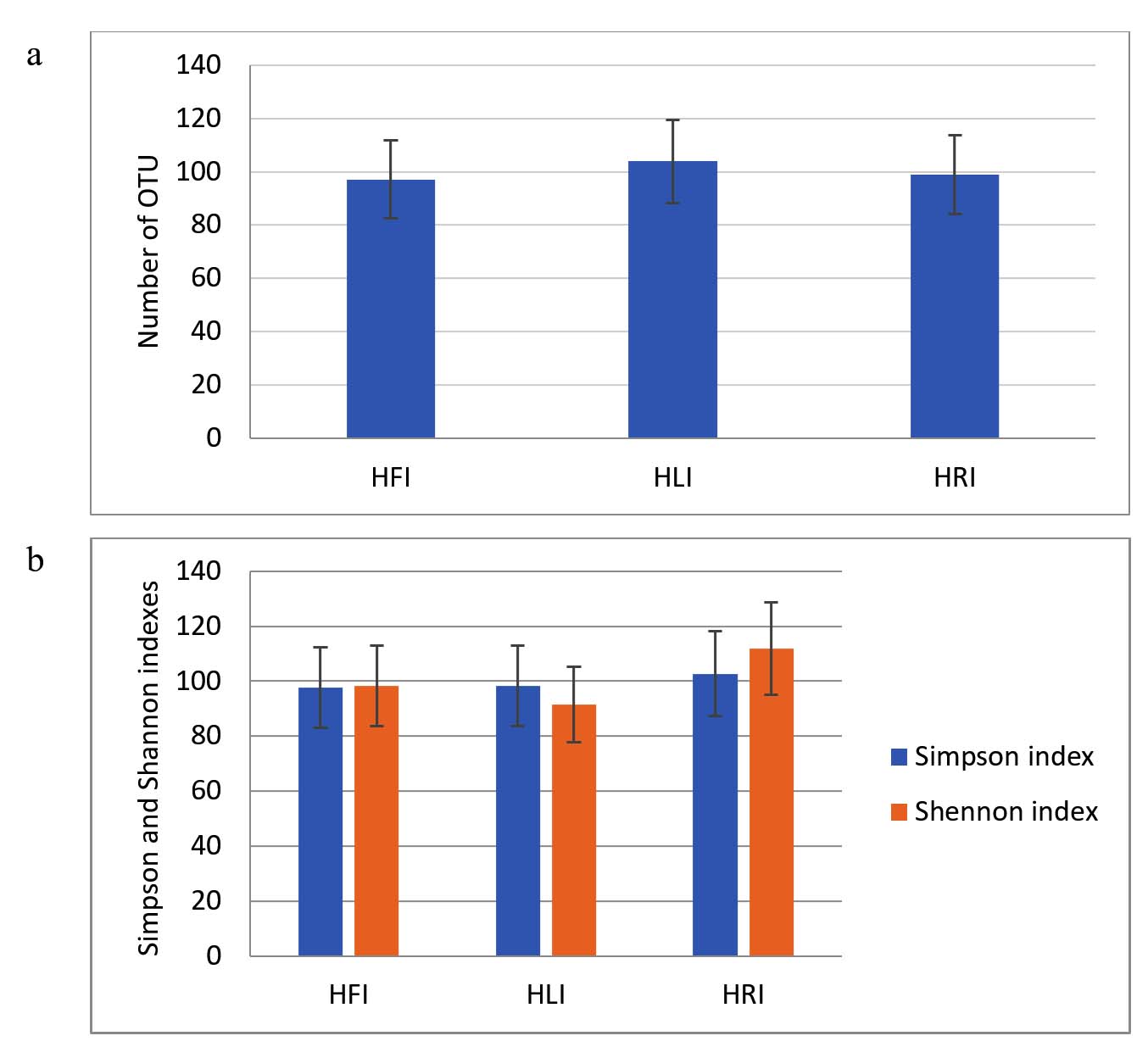
Figure 2. The number of OTUs (a), Simpson index, and Shannon index (b) in filtrate, leaves, and roots of hydroponically grown lettuce with IGSA chelated fertilizer addition
In the soil experiment control samples, the OTU count in soil was 1595, 3.5 times higher than in the control filtrate sample. In leaves, the OTU count was 44, not significantly different from the hydroponic experiment. In roots, the OTU count was 296, 1.7 times higher than in the hydroponic control roots. The Simpson and Shannon indices were 0.99 and 5.65 for soil, 0.75 and 1.97 for leaves, and 0.92 and 3.66 for roots. In the soil experiment, two types of IGSA treatments were used: foliar and root application. As with the hydroponic experiment, IGSA did not affect the alpha diversity of the microbial communities in soil, leaves, and roots, nor the OTU counts in samples, except for the root sample treated with IGSA by root application (sample SRIf), where the OTU count was 73.6% of the control (Figure 3).
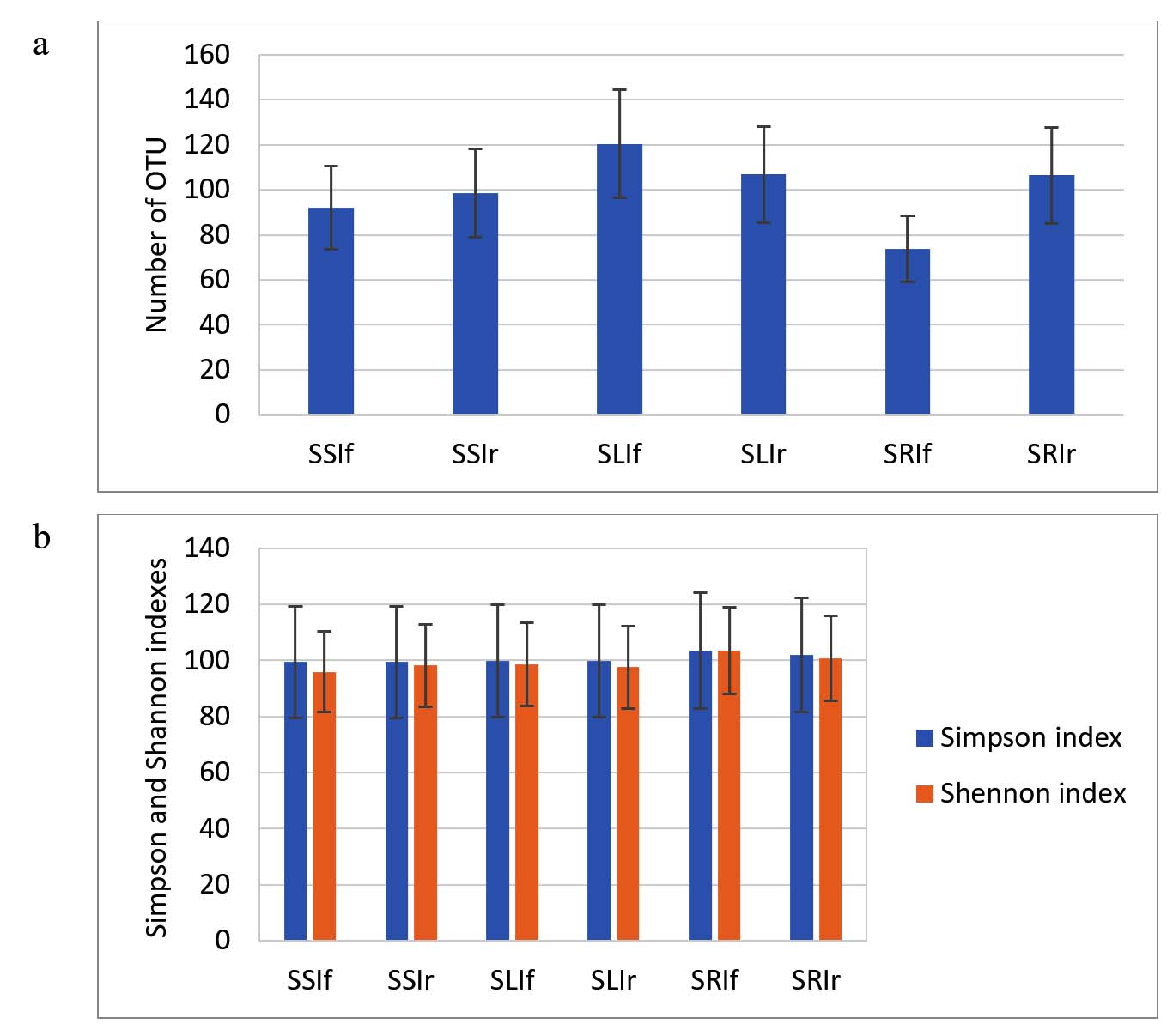
Figure 3. The number of OTUs (a), Simpson index, and Shannon index (b) in soil, leaves, and roots of lettuce grown in soil with IGSA chelated fertilizer addition
Thus, it was determined that in the substrates used for cultivating lettuce (soil, hydroponics), the number and alpha diversity of microorganisms were higher than in the roots and leaves of the plants. Soil contained significantly more microorganisms in both quantity and diversity compared to filtrate. Soil, unlike the aqueous medium, contains a wider range of soluble nutrients. Due to its structure and the presence of grains and aggregates of various sizes and textures, soil provides bacteria with a larger number of ecological niches. The higher number and diversity of microorganisms in the roots of soil-grown lettuce are likely due to some microorganisms being introduced from the soil environment. Bacterial penetration into root tissues can occur passively through root cracks or lateral root exit points, as well as through a number of active mechanisms.20,21 Unlike roots, no differences in the number of OTUs and alpha diversity indices were observed in the leaves of lettuce grown on different substrates (soil, hydroponics).
Next, the influence of IGSA on the composition and abundance of dominant bacterial taxa (OTU abundance over 2% in at least one sample) in hydroponic filtrate and soil of the soil experiment was evaluated (Table 1). The same bacterial taxa were present among the dominants, but their abundance varied depending on the substrate type. In the control hydroponic filtrate, the dominant bacteria were from the family Simkaniaceae (abundance 4.2) and the genera Glutamicibacter (15.02), Flavobacterium (5.5), Devosia (5.4), and Sphingomonas (6.7). In the control soil samples, the most frequently encountered bacteria were from the genera Pseudarthrobacter (abundance 7.2), Bacillus (2.9), and Sphingomonas (6.7).
Table (1): Dominant (OTU abundance over 2% in at least one sample) bacterial taxa in the microbial community of hydroponic filtrate and soil of the experiment with IGSA chelated fertilizer
| The lowest defined taxon | OTU | Hydroponics | Soil experiment | |||
|---|---|---|---|---|---|---|
| HFK | HFI | SSK | SSIr | SSIf | ||
| genus | Glutamicibacter | 15.0 | 0.3 | 0.2 | 0.2 | 0.2 |
| genus | Pseudarthrobacter | 2.1 | 19.1 | 7.2 | 10.1 | 8.8 |
| genus | Flavobacterium | 5.5 | 0.1 | 0.2 | 0.3 | 0.4 |
| family | Simkaniaceae | 4.2 | 2.9 | 0.2 | 0.2 | 0.3 |
| genus | Bacillus | 0.0 | 0.0 | 3.0 | 3.3 | 3.6 |
| family | Saccharimonadaceae | 2.9 | 0.8 | 0.2 | 0.3 | 0.2 |
| order | Saccharimonadales | 1.9 | 14.6 | 0.7 | 0.9 | 0.9 |
| genus | Brevundimonas | 2.2 | 1.3 | 0.4 | 0.2 | 0.5 |
| family | Paracaedibacteraceae | 3.1 | 0.0 | 0.0 | 0.0 | 0.0 |
| genus | Reyranella | 3.0 | 0.9 | 0.2 | 0.2 | 0.2 |
| genus | Devosia | 5.4 | 2.0 | 1.0 | 1.0 | 1.4 |
| genus | Allorhizobium-Neorhizobium- Pararhizobium-Rhizobium |
2.7 | 1.3 | 0.4 | 0.3 | 0.4 |
| family | Rhizobiaceae | 1.1 | 3.4 | 0.5 | 0.5 | 0.5 |
| family | Rickettsiaceae | 0.0 | 5.9 | 0.3 | 0.2 | 0.2 |
| genus | Novosphingobium | 2.2 | 0.2 | 0.2 | 0.3 | 0.4 |
| genus | Sphingobium | 3.7 | 1.1 | 0.3 | 0.4 | 0.2 |
| genus | Sphingomonas | 6.7 | 0.8 | 2.0 | 1.8 | 1.9 |
| genus | Limnobacter | 0.0 | 0.0 | 0.7 | 0.7 | 2.3 |
| genus | Methylophilus | 2.6 | 1.1 | 0.0 | 0.0 | 0.0 |
Adding IGSA to the hydroponic filtrate altered the abundance of most dominant bacterial taxa. An increase in the abundance of bacteria belonging to the genus Pseudarthrobacter, order Saccharimonadales, families Rhizobiaceae and Rickettsiaceae was observed. The abundance of bacteria from the genera Glutamicibacter, Flavobacterium, Reyranella, Devosia, and Sphingomonas, and the families Saccharimonadaceae and Paracaedibacteraceae decreased.
In the soil experiment, the effect of IGSA on the abundance of bacterial taxa was less pronounced compared to the hydroponic system. This difference is likely due to the filtrate being a more homogeneous medium compared to soil, which increases IGSA bioavailability. An increase in the abundance of bacteria belonging to the genus Pseudarthrobacter was observed in the IGSA-treated variants, both foliar and root applications (samples SSIr and SSIf).
Thus, the application of IGSA leads to changes in the abundance of some dominant strains in the substrate, particularly in hydroponic solutions. However, no elimination of any OTUs or the appearance of new dominants compared to the control occurred. The method of IGSA application (root/foliar) did not affect the composition and abundance of dominant bacterial taxa in soil and filtrate.
Soil serves as a reservoir for a large number and diversity of microorganisms compared to the aquatic environment. The number of bacteria in soil varies from less than 500,000 to 3,863,000 cells/mL, while in water, these numbers are significantly lower. Soil has a high buffering capacity, consistent pH, salinity, and other physicochemical properties. Furthermore, soil is not subject to rapid temperature changes and nutrient concentration fluctuations.22,23 In water, conditions for the microbial community are more variable. Fluctuations in salinity, thermal and oxygen stratification, and concentrations of ammonium, nitrates, total chlorine, and monochloramine can significantly affect the number of aquatic microorganisms.24,25 Nutrient deficiency limits microbial colonization of hydroponic systems. The number of microorganisms in it before planting is low, and the microbial community consists of a small number of species. Colonization of hydroponic systems by microorganisms mainly occurs through airborne droplets. For example, when cultivating plants using hydroponic systems, the filtrate can also be colonized by plant endophytes.26,27
Influence of IGSA on bacterial communities of Lettuce leaves and root endosphere
Next, the effect of IGSA chelated fertilizer on the endophytic microbial community of lettuce grown in soil and hydroponic systems was evaluated (Table 2). Differences in dominant bacterial taxa in the lettuce endophytic microbiome were observed depending on the substrate used (soil/hydroponics). When using soil for lettuce cultivation, bacteria Rhodococcus qingshengii sp. with an abundance of 11.5-34.6 were found in leaves and roots, absent in the endophytic microbiome of plants grown in hydroponics. Conversely, bacteria of the genus Methylophilus were present only in hydroponic experiment samples.
Table (2): Dominant (OTU abundance over 2% in at least one sample) bacterial taxa in the endophytic microbial community of lettuce grown in soil and hydroponic systems with IGSA chelated fertilizer
| The lowest defined taxon | OTU | Hydroponics | Soil experiment | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| leaf | root | leaf | root | ||||||||
| HLK | HLI | HRK | HRI | SLK | SLIr | SLIf | SRK | SRIr | SRIf | ||
| species | Rhodococcus qingshengii | 0.0 | 0.7 | 0.0 | 0.0 | 31.8 | 33.8 | 34.6 | 19.8 | 11.5 | 12.4 |
| genus | Pseudarthrobacter | 6.7 | 2.2 | 0.0 | 2.5 | 0.3 | 0.0 | 0.1 | 3.0 | 0.4 | 0.2 |
| genus | Streptomyces | 0.0 | 0.0 | 0.2 | 5.8 | 0.0 | 0.0 | 0.1 | 2.7 | 0.7 | 2.5 |
| family | Muribaculaceae | 0.7 | 11.4 | 0.3 | 0.0 | 0.0 | 0.0 | 0.1 | 0.0 | 0.7 | 1.2 |
| genus | Myroides | 0.0 | 0.0 | 0.0 | 0.0 | 0.9 | 0.3 | 2.4 | 0.7 | 0.2 | 0.6 |
| family | NS11-12 marine group | 2.7 | 0.9 | 0.0 | 1.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.1 |
| genus | Pedobacter | 2.2 | 0.7 | 0.4 | 0.5 | 0.0 | 0.0 | 0.0 | 0.2 | 0.0 | 0.2 |
| family | env.OPS 17 | 1.7 | 2.7 | 3.6 | 2.6 | 2.1 | 1.5 | 1.8 | 0.8 | 1.1 | 1.7 |
| family | Simkaniaceae | 5.3 | 1.8 | 0.5 | 0.1 | 0.0 | 0.0 | 0.0 | 2.9 | 6.5 | 5.5 |
| genus | Lactobacillus | 1.1 | 23.0 | 2.6 | 0.8 | 0.6 | 0.4 | 1.8 | 0.2 | 5.0 | 6.5 |
| genus | Candidatus Arthromitus | 0.0 | 2.4 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.6 | 1.0 |
| genus | Romboutsia | 0.0 | 0.2 | 3.6 | 0.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.4 | 0.5 |
| family | Peptostreptococcaceae | 0.0 | 0.1 | 3.2 | 0.2 | 0.0 | 0.1 | 0.1 | 0.0 | 0.6 | 0.3 |
| order | Saccharimonadales | 8.8 | 2.9 | 0.0 | 0.5 | 0.0 | 0.0 | 0.0 | 1.4 | 0.9 | 1.3 |
| genus | Reyranella | 4.1 | 2.4 | 0.8 | 1.8 | 0.8 | 0.9 | 0.0 | 0.4 | 0.3 | 0.3 |
| genus | Bosea | 1.7 | 0.6 | 0.0 | 2.3 | 0.0 | 0.0 | 0.0 | 0.2 | 0.0 | 0.1 |
| genus | Devosia | 2.1 | 0.7 | 0.1 | 1.2 | 0.0 | 0.1 | 0.0 | 2.5 | 0.5 | 0.7 |
| genus | Allorhizobium- Neorhizobium- Pararhizobium- Rhizobium | 1.1 | 0.4 | 6.7 | 2.5 | 0.0 | 0.0 | 0.0 | 1.0 | 4.2 | 0.8 |
| genus | Sphingobium | 1.9 | 0.6 | 0.6 | 2.6 | 0.0 | 0.0 | 0.0 | 0.4 | 0.2 | 0.0 |
| genus | Sphingomonas | 10.2 | 9.5 | 13.0 | 10.9 | 5.1 | 8.6 | 8.9 | 6.8 | 6.1 | 7.9 |
| genus | Achromobacter | 0.0 | 0.0 | 0.1 | 3.2 | 0.0 | 0.0 | 0.0 | 0.1 | 0.4 | 0.0 |
| genus | Aquabacterium | 1.4 | 1.4 | 0.6 | 1.4 | 4.1 | 3.6 | 4.9 | 2.1 | 2.1 | 2.8 |
| genus | Massilia | 0.8 | 0.3 | 9.9 | 2.4 | 0.0 | 0.0 | 0.0 | 0.6 | 0.0 | 0.5 |
| genus | Methylophilus | 1.0 | 0.3 | 18.6 | 19.7 | 0.0 | 0.0 | 0.0 | 0.0 | 0.2 | 0.2 |
| family | Methylophilaceae | 0.0 | 0.0 | 0.4 | 0.4 | 0.0 | 0.0 | 0.0 | 1.1 | 2.5 | 1.4 |
| family | Enterobacteriaceae | 0.0 | 2.7 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| genus | Pseudomonas | 0.0 | 0.4 | 5.1 | 1.7 | 37.5 | 35.0 | 30.8 | 13.5 | 12.5 | 6.9 |
| genus | Ureaplasma | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.1 | 0.2 | 0.0 | 2.6 | 2.5 |
In the control hydroponic lettuce leaf sample (HLK), the most abundant bacteria were from the genera Sphingomonas (abundance 10.2), Pseudarthrobacter (6.7), and Reyranella (4.1), family Simkaniaceae (5.3), and order Saccharimonadales (8.8). The endophytic microbial community of the root (HRK sample) was characterized by the following dominant genera: Allorhizobium-Neorhizobium-Pararhizobium-Rhizobium (abundance 6.7), Sphingomonas (13), Massilia (9.9), Methylophilus (18.6), and Pseudomonas (5.1).
Using IGSA in the hydroponic experiment increased the abundance of bacteria from the families Muribaculaceae and Enterobacteriaceae and the genus Lactobacillus in leaves. At the same time, the abundance of bacteria from the genus Pseudarthrobacter, family Simkaniaceae, and order Saccharimonadales decreased. In the root microbial community, an increase in the abundance of bacteria from the genera Pseudarthrobacter and Streptomyces and a decrease in the abundance of bacteria from the families Peptostreptococcaceae and genera Romboutsia, Massilia, Pseudomonas, and Allorhizobium–Neorhizobium–Pararhizobium–Rhizobium occurred.
In the control soil-grown lettuce leaf sample, the dominant bacterial taxa were Rhodococcus qingshengii sp. (abundance 31.8) and genus Pseudomonas (37.5). In the root sample, the most frequent bacteria were Rhodococcus qingshengii sp. (abundance 19.8), genera Sphingomonas (6.8), and Pseudomonas (13.5). With the application of chelated fertilizer by both methods (root/foliar), an increase in the abundance of bacteria from the genus Sphingomonas and a slight decrease in the abundance of bacteria from the genus Pseudomonas were observed in the endophytic microbial community of lettuce leaves. Interestingly, using IGSA for lettuce grown in soil had a less pronounced effect on the number of dominant endophytic bacteria compared to its cultivation in hydroponics. The heterogeneous soil structure and sorption processes could have reduced the bioavailability of IGSA. Moreover, assembling the root microbiome in hydroponics fundamentally differs from that in soil, where plants can attract microbial partners from a more diverse pool. This assembly method leads to greater stability of the root microbiome in plants grown in soil compared to hydroponics.26,28
Overall, the results obtained for the endospheric communities of lettuce are similar to those observed for substrates: the application of chelated fertilizer led to some changes in species composition and diversity, mainly when grown in hydroponics. However, no drastic changes in communities from the original (control) state occurred. The method of IGSA application did not lead to any additional changes in community composition.
Core endophytic microbiome of lettuce plants
Next, we identified bacterial taxa that constitute the core endophytic microbial community of lettuce leaves and roots, irrespective of the growing conditions (hydroponics/soil) (Figure 4). The core microbiome of plants consists of key microorganisms that contain genes selected through evolutionary mechanisms and necessary for plant function and adaptation.29,30
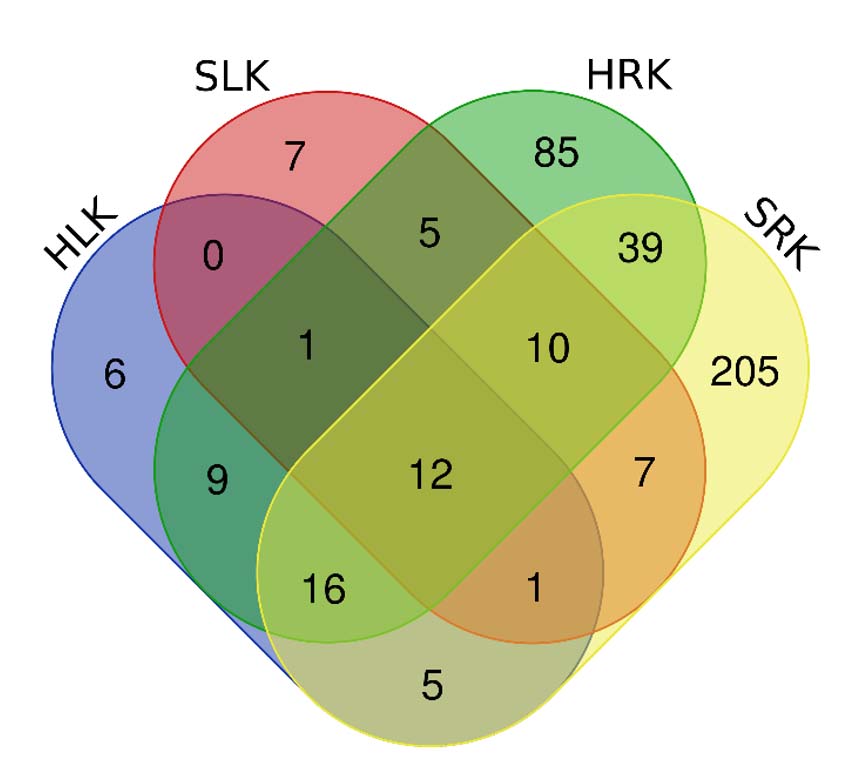
Figure 4. Overlapping bacterial OTUs in control samples of lettuce leaves and roots grown in soil and hydroponics
The core microorganisms for lettuce included 12 bacterial OTUs (Table 3). These microorganisms can inhabit various plants and many other ecological niches (soil, water, biofilms, aquifers, lake sediments).31-34 Additionally, bacteria from some detected taxa (genus Ralstonia and genus Sphingomonas) are plant pathogens.35-38
Table (3): Core bacterial OTUs in control samples of lettuce leaves and roots grown in soil and hydroponics
The lowest defined taxon |
OTU |
|---|---|
order |
Obscuribacterales |
genus |
Bradyrhizobium |
genus |
Reyranella |
family |
env.OPS 17 |
genus |
Ralstonia |
family |
Caulobacteraceae |
genus |
Sphingomonas |
genus |
Pelomonas |
genus |
Methylobacterium |
genus |
Aquabacterium |
genus |
Sphingomonas |
genus |
Lactobacillus |
Next, we identified unique OTUs present in control samples of lettuce leaves and roots depending on the substrate used. In lettuce grown in hydroponics, there were 6 and 85 unique OTUs in leaves and roots, respectively. When using soil, 7 and 205 unique bacterial OTUs were found in leaves and roots, respectively. Interestingly, using different substrates did not affect the number of unique OTUs in leaves, but significant differences were found in the number of unique OTUs in lettuce roots. Root-associated microorganisms mainly come from soil. The rhizosphere microbial community strongly influences the root microbiome composition. Soil microbial community structure and environmental parameters, including the substrate, are considered more critical factors for root bacterial community structure than plant genotype or species.39,40
Next, the microbial communities of lettuce leaves and roots grown on different substrates with and without IGSA chelated fertilizer were analyzed separately (Figure 5 a, b). Among the leaf microbial community, 9 core microorganisms were found regardless of the substrate and fertilizer treatment. More unique taxa (30 OTUs and 23 OTUs) were found in lettuce leaf samples grown in hydroponics, regardless of IGSA treatment (samples HLK and HLI). Overall, comparing microbial communities in root samples revealed more core and unique OTUs than in leaf samples. For example, 36 root core OTUs were found, three times the number of core OTUs in leaf samples. The analysis of core OTUs in microbial communities revealed 8 common endophytes for leaves and roots, present in lettuce plants regardless of growing conditions and IGSA treatment. Notably, the most significant differences in the number of unique OTUs were found between different root samples rather than leaf samples.
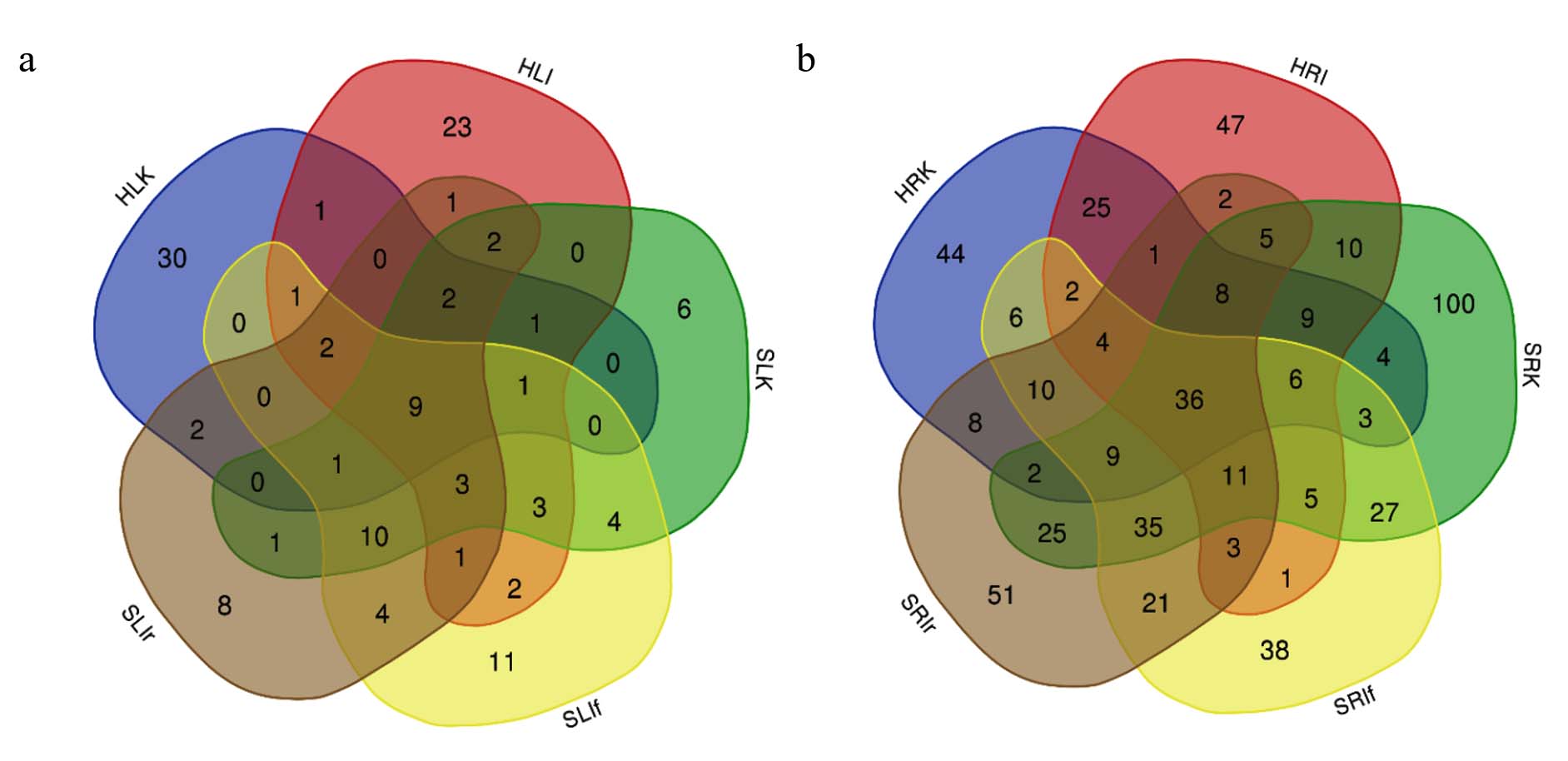
Figure 5. Overlapping bacterial OTUs in lettuce leaves (a) and root (b) samples grown in soil and hydroponics with IGSA chelated fertilizer
Beta-diversity of bacterial communities of soil, hydroponics, and Lettuce plants endosphere
Finally, non-metric multidimensional scaling (NMDS) was used to analyze the similarities and differences in microbial communities of lettuce leaves and roots, soil, and hydroponics. In Figure 6, each point represents an individual sample, and a greater distance between them indicates greater differences between the communities of the respective samples. The figure shows that points representing endophytic communities and substrate communities can be divided into two groups, H and S by substrate type. Within the S group, three subgroups of points corresponding to soil, leaf, and root communities can be distinguished.
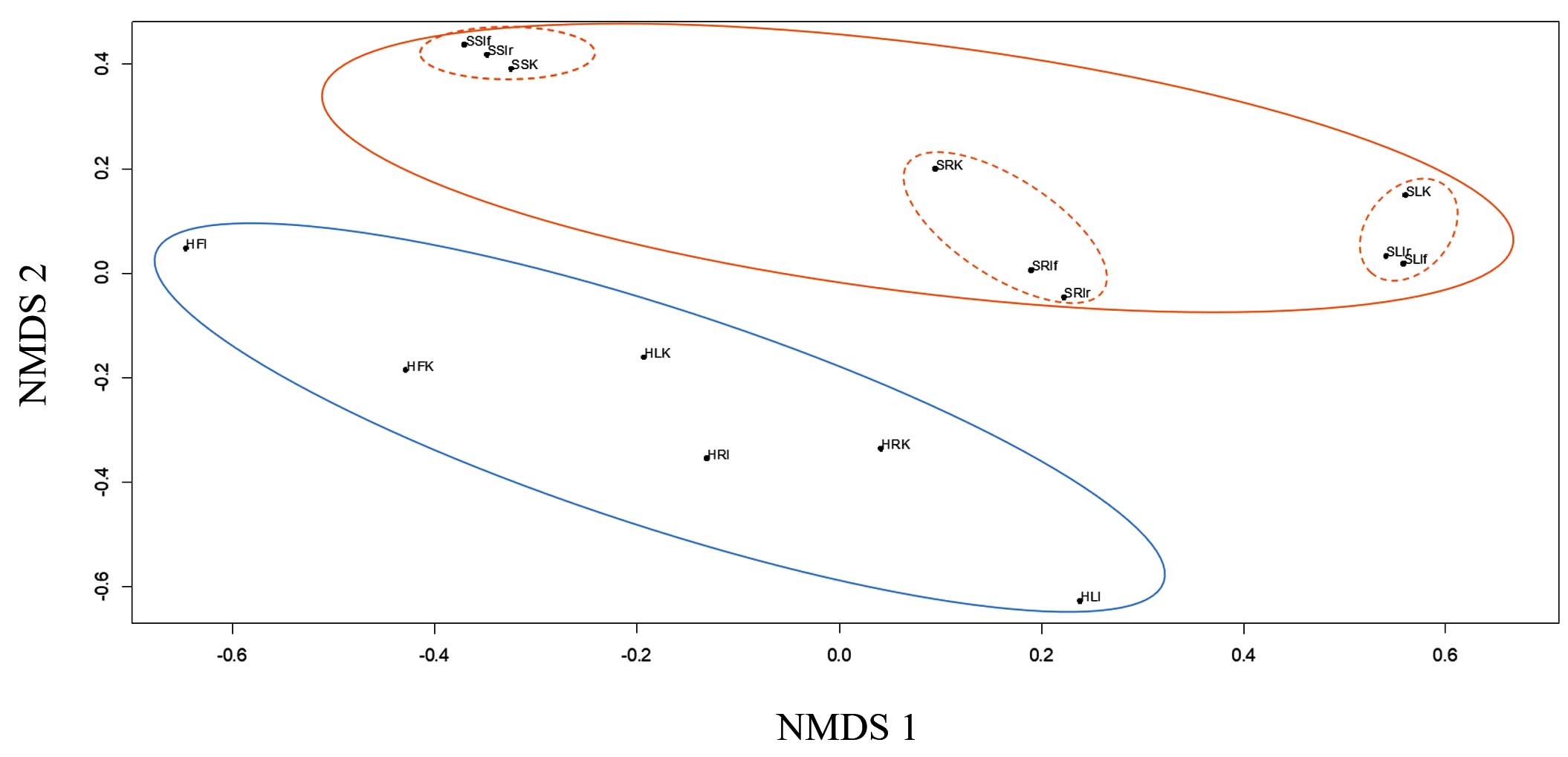
Figure 6. NMDS analysis of the bacterial communities of filtrate, soil and lettuce without and with IGSA chelated fertilizer
Within each subgroup, distances between points representing communities in control and IGSA-treated variants do not differ. NMDS analysis confirms the data obtained using other sequencing analysis methods: the substrate type (soil, hydroponics) has the most significant influence on forming endophytic communities. Chelate application does not affect the composition of endophytic or substrate communities. Lettuce root and leaf communities differ significantly, especially when grown in soil, likely due to rhizosphere microorganisms migrating into plant tissues.
This study demonstrates that the new IGSA chelating micronutrient fertilizer does not adversely affect the endophytic microbial community of lettuce. The substrate type remains the most significant factor influencing microbial community composition and diversity. The findings support the potential use of IGSA as a safe and effective alternative to traditional EDTA-based fertilizers, contributing to more sustainable agricultural practices. The results suggest that the use of IGSA as a chelating agent in micronutrient fertilizers is a promising and environmentally safe alternative to EDTA. Given the minimal impact of IGSA on the endophytic microbial communities of lettuce, it could be recommended for broader agricultural use. The study highlights the importance of considering substrate types in agricultural practices, as they significantly influence microbial communities associated with plants. Future research should explore the long-term effects of IGSA on various crops and in different environmental conditions. Additionally, understanding the mechanisms behind the interaction between IGSA and plant-associated microorganisms will provide deeper insights into optimizing fertilizer use for sustainable agriculture.
Additional file: Table S1.
ACKNOWLEDGMENTS
None.
CONFLICT OF INTEREST
The authors declare that there is no conflict of interest.
AUTHORS’ CONTRIBUTION
ND, GG and PG conceptualized the study and applied methodology. ND, LB, PK and GG performed Investigation, material preparation, data collection and formal analysis. ND and GG wrote the manuscript. SS performed supervision and project administration. All authors read and approved the final manuscript for publication.
FUNDING
This study was funded by the Kazan Federal University for the state assignment in the sphere of scientific activities, project number FZSM-2026-0029.
DATA AVAILABILITY
All datasets generated or analyzed during this study are included in the manuscript and/or in the supplementary files.
ETHICS STATEMENT
Not applicable.
- Ren Z. Effects of risk perception and agricultural socialized services on farmers’ organic fertilizer application behavior: Evidence from Shandong Province, China. Front Public Heal. 2023;11(15):1-11.
Crossref - Beltyukova M, Kuryntseva P, Galitskaya P, Selivanovskaya S, Brusko V, Dimiev A. Biodegradation rate of edta and ids and their metal complexes. Horticulturae. 2023;9(6):1-10.
Crossref - Guo JK, Lv X, Jia HL, et al. Effects of EDTA and plant growth-promoting rhizobacteria on plant growth and heavy metal uptake of hyperaccumulator Sedum alfredii Hance. J Environ Sci. 2020;88(15):361-369.
Crossref - Gluhar S, Kaurin A, Lestan D. Soil washing with biodegradable chelating agents and EDTA: Technological feasibility, remediation efficiency and environmental sustainability. Chemosphere. 2020;257(12):127226.
Crossref - Huang R, Cui X, Luo X, et al. Effects of plant growth regulator and chelating agent on the phytoextraction of heavy metals by Pfaffia glomerata and on the soil microbial community. Environ Pollut. 2021;283(3):117159.
Crossref - Brusko V, Garifullin B, Geniyatullina G, et al. Novel biodegradable chelating agents for micronutrient fertilization. J Agric Food Chem. 2023;71(41):14979-14988.
Crossref - Liu X, Zhu X, Dong Y, Chen Y, Li M, Li C. Limited impact of soil microorganisms on the endophytic bacteria of Tartary buckwheat (Fagopyrum tataricum). Microorganisms. 2023;11(8):1-15.
Crossref - Miranda-Carrazco A, Navarro-Noya YE, Govaerts B, Verhulst N, Dendooven L. Nitrogen fertilizer application alters the root endophyte bacterial microbiome in maize plants, but not in the stem or rhizosphere soil. Microbiol Spectr. 2022;10(6):1-14.
Crossref - Liu Y, Chu G, Stirling E, et al. Nitrogen fertilization modulates rice seed endophytic microbiomes and grain quality. Sci Total Environ. 2023;857(part 2):159181.
Crossref - Wippel K. Plant and microbial features governing an endophytic lifestyle. Curr Opin Plant Biol. 2023;76(2):102483.
Crossref - Caporaso JG, Kuczynski J, Stombaugh J, et al. QIIME allows analysis of high-throughput community sequencing data. Nat Methods. 2010;7(5):335-336.
Crossref - Wang Q, Garrity GM, Tiedje JM, Cole JR. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl Environ Microbiol. 2007;73(16):5261-5267.
Crossref - Clarke KR. Non parametric multivariate analyses of changes in community structure. Aust J Ecol. 1993;18(1):117-143.
Crossref - Simpson EH. Measurment of Diversity. Nature. 1949;163(4148):688.
Crossref - Garvey RS, Fowler AC. On the mathematical theory of plumes. Geophys Astrophys Fluid Dyn. 2023;117(2):79-106.
Crossref - Tudi M, Ruan HD, Wang L, et al. Agriculture development, pesticide application and its impact on the environment. Int J Environ Res Public Health. 2021;18(3):1-24.
Crossref - Shadab M, Bhatti N, Ain Q, et al. Allelopathy for the sustainable management of agricultural pests: Appraisal of major allelochemicals and mechanisms underlying their actions. S Afr J Bot. 2024;175(5):496-513.
Crossref - Chen X, Henriksen TM, Svensson K, Korsaeth A. Long-term effects of agricultural production systems on structure and function of the soil microbial community. Appl Soil Ecol. 2020;147(9):103387.
Crossref - Zhang Y, Zhang H, Tao T, et al. Investigation and insights into the technical strategies for reducing agrochemical inputs in rice farming to enhance agroecosystem resilience and food safety, A case study in China. J Environ Manage. 2025;392(7):126619.
Crossref - Mhete M, Eze PN, Rahube TO, Akinyemi FO. Soil properties influence bacterial abundance and diversity under different land-use regimes in semi-arid environments. Sci African. 2020;7(11):1-13.
Crossref - De Carvalho-Niebel F, Fournier J, Becker A, Arancibia MM. Cellular insights into legume root infection by rhizobia. Curr Opin Plant Biol. 2024;81:102597.
Crossref - Caillon F, Schelker J. Dynamic transfer of soil bacteria and dissolved organic carbon into small streams during hydrological events. Aquat Sci. 2020;82(2):1-11.
Crossref - Iqbal S, Begum F, Nguchu BA, Claver UP, Shaw P. The invisible architects: microbial communities and their transformative role in soil health and global climate changes. Environ Microbiome. 2025;20(1):36.
Crossref - Kelly JJ, Minalt N, Culotti A, Pryor M, Packman A. Temporal variations in the abundance and composition of biofilm communities colonizing drinking water distribution pipes. PLoS One. 2014;9(5):1-12.
Crossref - Hu W, Zhou X, Liu Y, Zhang Y, Wang Y. Effects of salinity fluctuation on antimicrobial resistance and virulence factor genes of low and high nucleic acid-content bacteria in a marine environment. Microorganisms. 2025;13(7):1-16.
Crossref - Thomas P, Knox OGG, Powell JR, Sindel B, Winter G. The Hydroponic rockwool root microbiome: under control or underutilised? Microorganisms. 2023;11(4):1-17.
Crossref - Thomas BO, Lechner SL, Ross HC, Joris BR, Glick BR, Stegelmeier AA. Friends and foes: bacteria of the hydroponic plant microbiome. Plants. 2024;13(21):1-32.
Crossref - Kuryntseva P, Pronovich N, Galieva G, Galitskaya P, Selivanovskaya S. Exploring the role of vertical and horizontal pathways in the formation of lettuce plant endospheric bacterial communities: a comparative study of hydroponic and soil systems. Horticulturae. 2025;11(7):1-22.
Crossref - Compant S, Samad A, Faist H, Sessitsch A. A review on the plant microbiome: Ecology, functions, and emerging trends in microbial application. J Adv Res. 2019;19(5):29-37.
Crossref - Duran P. The core microbiota across the green lineage. Curr Opin Plant Biol. 2024;77:102487.
Crossref - Ruiz-Gil T, Acuna JJ, Fujiyoshi S, et al. Airborne bacterial communities of outdoor environments and their associated influencing factors. Environ Int. 2020;145(6):1-14.
Crossref - Wang Z, Liu J, Xu H, Liu J, Zhao Z, Gong X. Core microbiome and microbial community structure in Coralloid Roots of Cycas in Ex Situ Collection of Kunming Botanical Garden in China. Microorganisms. 2023;11(9):1-18.
Crossref - Lee H, Kim DU, Lee S, et al. Reyranella terrae sp. Nov., isolated from an agricultural soil, and emended description of the genus Reyranella. Int J Syst Evol Microbiol. 2017;67(6):2031-2035.
Crossref - Procter M, Kundu B, Sudalaimuthuasari N, et al. Microbiome of Citrullus colocynthis (L.) Schrad. reveals a potential association with non-photosynthetic Cyanobacteria. Microorganisms. 2022;10(10):1-13.
Crossref - Yanqing L, Sun W, Chen X, et al. Bacterial Leaf Spot of Plum Caused by Sphingomonas spermidinifaciens in Guangxi, China. Plant disease. 2023;107(9): 2839.
Crossref - Pratap SD, Ramesh R, Maurya S, et al. Metabolic reprogramming of tomato plants under Ralstonia solanacearum infection. Plant Stress. 2025;16(12):100804.
Crossref - Parihar M, Rakshit A, Meena VS, et al. The potential of arbuscular mycorrhizal fungi in C cycling: a review. Arch Microbiol. 2020;202(7):1581-1596.
Crossref - Yuan T, Yi H, Huang X, Li R, Qazi IH, Liu J. Understanding plant bacterial wilt: pathogen diversity, ecological interactions, and management challenges. Ecol Indic. 2025;181:114423.
Crossref - Dastogeer KMG, Tumpa FH, Sultana A, Akter MA, Chakraborty A. Plant microbiome–an account of the factors that shape community composition and diversity. Curr Plant Biol. 2020;23:100161.
Crossref - Debbagh-nour H, Khourchi S, et al. Plant identity and environmental filtering are the key drivers of bacterial community structure in four desert plant species from the Sahara Desert in Morocco. BMC Microbiol. 2025;25:1-16.
Crossref
© The Author(s) 2026. Open Access. This article is distributed under the terms of the Creative Commons Attribution 4.0 International License which permits unrestricted use, sharing, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.