


ISSN: 0973-7510
E-ISSN: 2581-690X
The emergence of multidrug-resistant Klebsiella pneumoniae is a major obstacle in the treatment of urinary tract infections (UTIs), and integron-mediated gene transfer contributes to this problem. This cross-sectional study included K. pneumoniae isolates obtained from urine samples of 200 patients diagnosed with UTI. Isolates were identified using culture and biochemical tests, and antimicrobial susceptibility was determined by disc diffusion against 14 antibiotics, including ampicillin, amoxicillin, cefotaxime, ceftriaxone, cephalexin, tetracycline, chloramphenicol, nalidixic acid, trimethoprim, ciprofloxacin, gentamicin, amikacin, and imipenem according to CLSI guidelines. Detection of class 1 integron integrase gene intI1 and gene cassettes dfrA5, dfrA7, and dfrA25 was performed by polymerase chain reaction using gene-specific primers that amplify target DNA sequences. Multidrug-resistance was observed in 94% of isolates, with high resistance to cephalexin (100%), ampicillin (99%), amoxicillin (99.5%), and cefotaxime (93%), whereas lower resistance was noted to imipenem (17%), gentamicin (21%), amikacin (21.5%), and ciprofloxacin (26%) (P < 0.05). PCR analysis showed that 31% of isolates carried the class 1 integron gene, while dfrA5, dfrA7, and dfrA25 were detected in 10%, 17%, and 12% of isolates, respectively, with dfrA7 being the most frequent cassette associated with integron-positive isolates. These findings indicate a high burden of multidrug-resistant K. pneumoniae in UTIs and highlight the role of class 1 integrons in the dissemination of resistance genes, emphasizing the need for continuous molecular surveillance and rational antibiotic use.
K. pneumoniae, UTIs, Resistance Patterns, Antibiotics, IntI1 gene
Urinary tract infection (UTI) is one of the most common bacterial infections in both community and hospital settings.1 Although Escherichia coli accounts for nearly 80% of UTIs, Klebsiella pneumoniae is an important secondary pathogen associated with recurrent and complicated infections.2 The increasing prevalence of antimicrobial-resistant K. pneumoniae has made empirical therapy more difficult, posing a serious challenge for clinicians.3 Excessive and inappropriate antibiotic use, together with horizontal gene transfer mediated by mobile genetic elements such as plasmids, transposons, and integrons, has accelerated the spread of multidrug-resistance among Enterobacteriaceae.4
Integrons are genetic platforms that capture and express antibiotic resistance genes through site specific recombination. They typically consist of an integrase gene (intI), an attachment site, and a promoter that drives expression of integrated gene cassettes.4,5 Among different integron classes, class 1 integrons are the most prevalent in clinical Gram-negative bacteria and contain variable regions with resistance gene cassettes located between conserved 5′ and 3′ segments.6-9 These gene cassettes encode resistance to multiple antibiotic classes, and their expression depends on the integron promoter.10,11 More than 130 gene cassettes have been described, including members of the aadA and dfrA families that confer resistance to streptomycin and trimethoprim, respectively.12,13
Although integrons and their gene cassettes have been widely reported in E. coli, Pseudomonas aeruginosa, and Acinetobacter baumannii isolates,14 information on their distribution in uropathogenic K. pneumoniae from Iraq remains limited.15 Increasing rates of recurrent UTIs and treatment failure in Dhi Qar city highlight the need for local molecular surveillance. Therefore, this study aimed to determine the prevalence of class 1 integrons and selected resistance gene cassettes and to evaluate their association with antibiotic resistance patterns in clinical K. pneumoniae isolates from patients with UTIs in northern Dhi Qar, Iraq.
Sample collection
This cross-sectional study was conducted from December 1, 2023, to March 24, 2024, at Al-Rifai General Hospital, Dhi Qar, Iraq. Urine samples were obtained once from 200 patients clinically suspected of urinary tract infection and confirmed by laboratory diagnosis. Diagnosis was based on compatible clinical symptoms (dysuria, urgency, frequency, fever, chills, or flank pain) together with significant bacteriuria (≥105 CFU/mL of a single predominant organism), considering patient age and clinical context.
Patients receiving antibiotics prescribed by their physicians were included, and individuals with other chronic diseases or concurrent bacterial infections were excluded. Written informed consent was obtained from all participants before sample collection. Midstream clean catch urine samples were collected from toilet trained patients in sterile screw cap containers, whereas urine from infants under two years of age was collected using sterile adhesive urine collection bags.7
Identification of bacterial isolates
Urine samples were cultured immediately on blood agar and MacConkey agar plates to isolate Enterobacteriaceae and assess lactose fermentation. Plates were incubated aerobically at 37 °C for 24 hrs, and colonies were examined for morphology, pigmentation, and hemolysis. Representative lactose-fermenting colonies suggestive of K. pneumoniae were subcultured on nutrient agar to obtain pure isolates. Identification was confirmed using standard biochemical tests in accordance with established microbiological procedures.2,16
Antimicrobial susceptibility testing
Antimicrobial susceptibility of confirmed uropathogenic K. pneumoniae isolates was determined by the Kirby-Bauer disc diffusion method on Mueller-Hinton agar following Clinical and Laboratory Standards Institute (CLSI) guidelines. Antibiotic discs representing drugs routinely tested for Enterobacteriaceae were applied as listed in Table 1, and plates were incubated at 37 °C for 18-24 hrs. Zone diameters were measured and interpreted as susceptible, intermediate, or resistant according to CLSI breakpoints.2,16
Table (1): Primers sequences and PCR annealing temperature of dfrA5, dfrA7 & dfrA25
| Genes | Primers sequences (5′-3′) | Annealing | Molecular weight |
|---|---|---|---|
| dfrA5 | Forward: GCCGGAATTCTAACCCAGGACGAGTAC | 56 °C for 30 sec | 510 bp |
| Reverse: GTCGAATTCTGACCCAGGAACGAGTTTC | |||
| dfrA7 | Forward: TCGAATTCTAACCCAGGACGAGTGGGC | 54 °C for 35 sec | 277 bp |
| Reverse: GTCGAATTCTGGGCGCGTTAGTTCCCCG | |||
| dfrA25 | Forward: CTGGAATTGGTAACCCAGGAAGCGCAC | 50 °C for 35 sec | 770 bp |
| Reverse: TCGAAATCTGGGTCCAGGAACGAGTGCC |
DNA extraction
Genomic DNA was extracted from confirmed Klebsiella pneumoniae isolates using a modified boiling method.16 Briefly, 300 µL of bacterial suspension adjusted to 0.5 McFarland turbidity was mixed with sterile distilled water in a 1.5 mL microcentrifuge tube, vortexed, and heated at 100 °C for 10 min in a water bath. The lysate was centrifuged at 14,000 × g for 10 min, and the clear supernatant containing genomic DNA was collected and stored at -20 °C until PCR analysis.
PCR for integron gene detection
The presence of the class 1 integron integrase gene intI1 was detected by polymerase chain reaction using primers F: AGTGGGTGGCGAATGAGTG and R: TGTTCTTGTATCGGCAGGTG.16 Each 25 µL reaction mixture contained 12.5 µL of 2× PCR Master Mix (Amplicon, Denmark), 0.3 µL of each primer (20 pmol/µL), 2.5 µL of template DNA, and nuclease free water to volume. Amplification was performed in a thermal cycler with initial denaturation at 94 °C for 4 min, followed by 35 cycles of denaturation at 94 °C for 40 sec, annealing at 58 °C for 30 sec, and extension at 72 °C for 40 sec, with a final extension at 72 °C for 5 min. PCR products were separated by electrophoresis on a 1.5% agarose gel stained with ethidium bromide and visualised under UV illumination. The presence of an amplicon of expected size confirmed detection of the intI1 gene.
PCR for gene cassette array detection
Detection of trimethoprim resistance gene cassettes dfrA5, dfrA7, and dfrA25 was performed by polymerase chain reaction using gene specific primers designed in this study. PCR assays were carried out using genomic DNA from confirmed K. pneumoniae isolates, particularly those positive for the class 1 integron-integrase gene (intI1). Reaction mixtures and cycling conditions were identical to those used for intI1 amplification, except for gene specific annealing temperatures as listed in Table 1. Amplified products were analyzed by agarose gel electrophoresis, and the presence of bands of expected size was considered positive for the respective gene cassette.
Statistical analysis
Data were analyzed using Microsoft Excel 2010 and SPSS version 19. Categorical variables were compared using the chi-square test or Fisher exact test when appropriate. Continuous variables were analyzed using the Mann-Whitney U test. A P-value less than 0.05 was considered statistically significant.
K. pneumoniae isolates were recovered from urine samples by culture and confirmed by biochemical testing. A total of 200 patients aged 1-65 years were included, with a mean age of 32.7 ± 11.3 years. UTIs were significantly more common in females than males (81% vs 19%, P = 0.006) as shown in Table 2. Age group analysis revealed a higher frequency of infection among patients aged 31-40 years (31%) and 51-65 years (24%), whereas the lowest prevalence was observed in children upto 10 years (6%) (Figure 1).
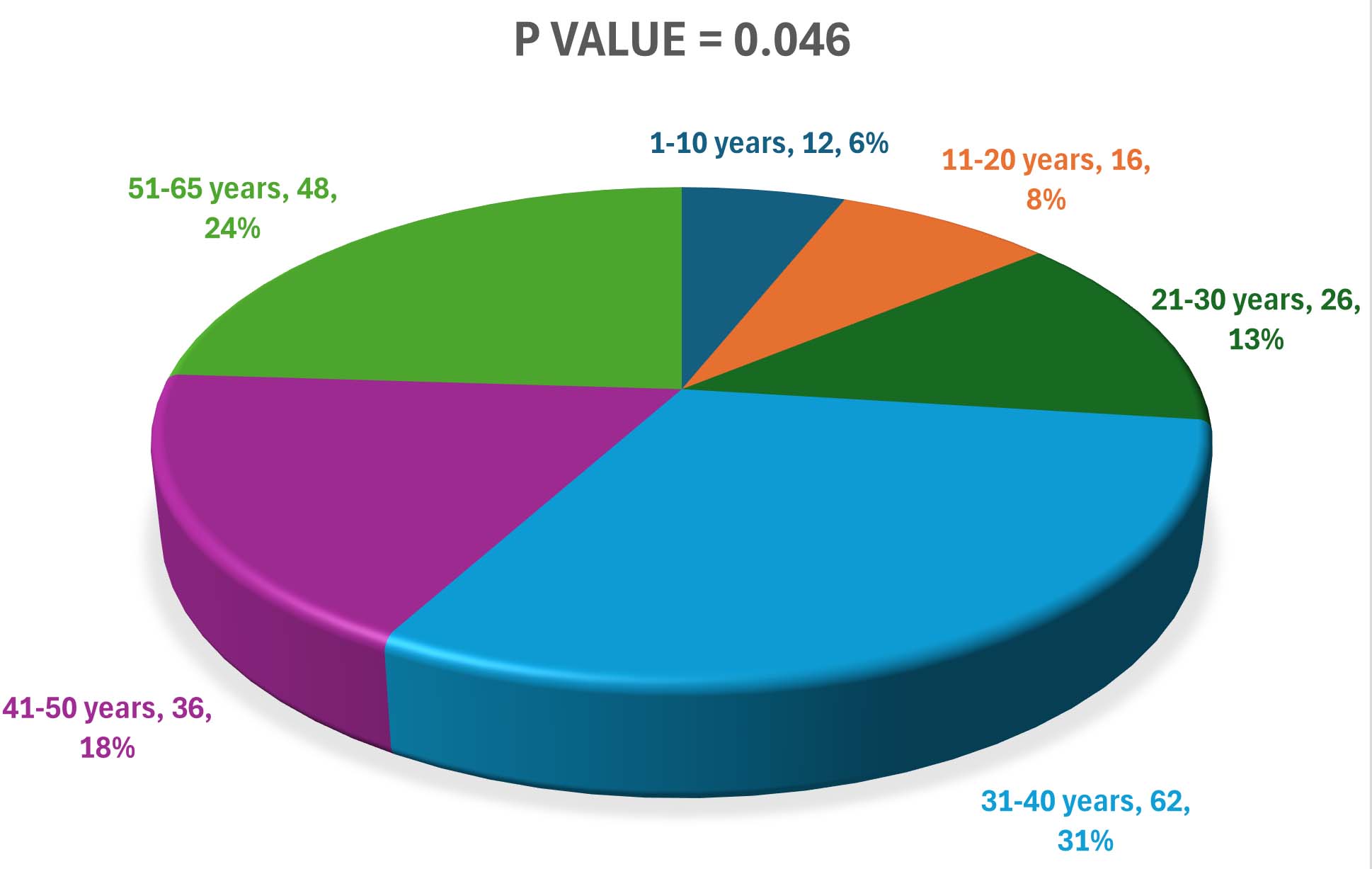
Figure 1. Frequency of K. pneumoniae infection according to patients age groups
Table (2): Age and gender properties of UTI patients
Character |
Data |
|---|---|
Age range |
1-65 years |
Age mean |
32.7 |
Standard deviation |
± 11.3 |
Standard error |
0.799 |
Gender |
N (%) |
Females |
162 (81%)* |
Males |
38 (19%) |
Total number |
200 |
*P-value (0.006) compared with males
Antimicrobial susceptibility patterns of K. pneumoniae isolates determined by disc diffusion are presented in Figure 2 and Table 3. Significant differences were observed between resistance and susceptibility rates among the tested antibiotics (P < 0.05). High resistance was detected to commonly used antibiotics, particularly cephalexin (100%), ampicillin (99%), amoxicillin (99.5%), and cefotaxime (93%). In contrast, lower resistance rates were observed for imipenem (17%), gentamicin (21%), amikacin (21.5%), and ciprofloxacin (26%). Overall, 94% of isolates were classified as multidrug-resistant.
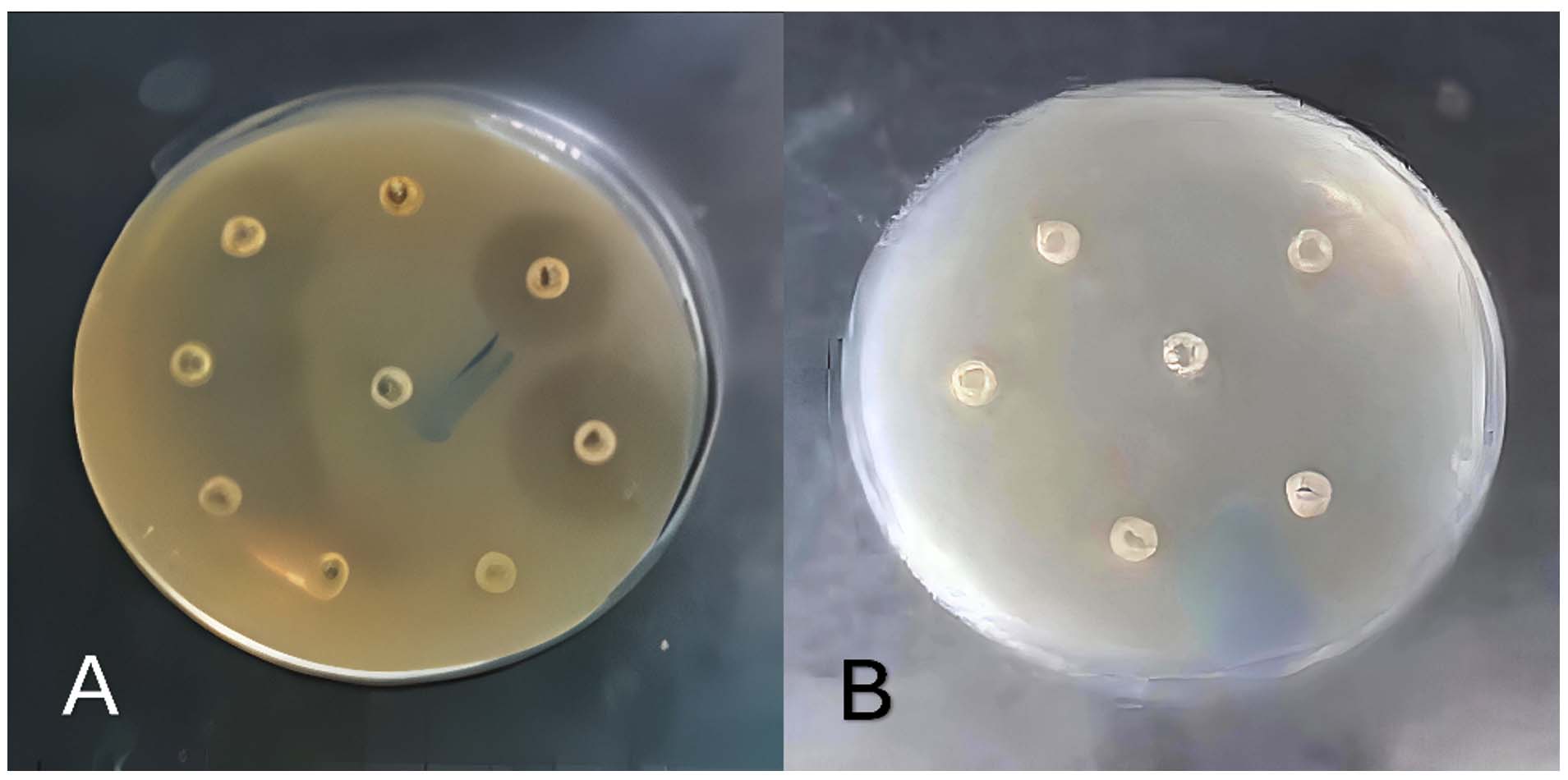
Figure 2. (A) Sensitivity of some isolates to antibiotics; (B) Resistance of some isolates to antibiotics
Table (3): Results of antibiotic sensitivity test of uropathogenic Klebsiella pneumoniae
Antibiotic agents |
Resistance N (%) |
Sensitivity N (%) |
χ2 |
P-value |
|---|---|---|---|---|
Chloramphenicol |
72 (36) |
128 (64) |
7.53 |
0.012 |
Tetracycline |
180 (90) |
20 (10) |
13.8 |
0.0018 |
Lincomycin |
134 (67) |
66 (33) |
17.76 |
0.010 |
Ampicillin |
198 (99) |
2 (1) |
26.1 |
|
Amoxicillin |
199 (99.5) |
1 (0.5) |
29.7 |
|
Cefotaxime |
186 (93) |
14 (7) |
13.14 |
0.0001 |
Ceftriaxone |
98 (48) |
72 (36) |
12.4 |
0.021 |
Cephalexin |
200 (100) |
0 (0) |
0.075 |
0.61 |
Nalidixic acid |
108 (54) |
92 (46) |
1.08 |
0.111 |
Trimethoprim |
102 (51) |
98 (49) |
0.035 |
0.822 |
Ciprofloxacin |
52 (26) |
148 (74) |
9.40 |
0.029 |
Amikacin |
43 (21.5) |
158 (78.5) |
14.1 |
0.030 |
Gentamicin |
42 (21) |
158 (79) |
13.6 |
0.028 |
Imipenem |
34 (17) |
166 (83) |
11.94 |
0.005 |
MDR |
188 (94) |
12 (6) |
15.85 |
PCR analysis (Figure 3) showed that 31% of K. pneumoniae isolates carried the class 1 integron-integrase gene (intI1). The trimethoprim resistance gene cassettes dfrA5, dfrA7, and dfrA25 were detected at lower frequencies of 10%, 17%, and 12%, respectively (Table 4). Eleven isolates harbored class 1 integrons containing multiple cassette genes (intI1-dfrA5-dfrA7-dfrA25), and dfrA7 was the most frequently detected cassette among integron-positive isolates (20 isolates). Overall, most isolates lacked these cassette genes (P < 0.05). Comparison of antibiotic resistance patterns according to gene presence showed no statistically significant differences (P > 0.05), although isolates carrying intI1 tended to exhibit higher resistance rates. No ciprofloxacin resistance was observed among isolates carrying dfrA5 or dfrA7.

Figure 3. PCR products of primers amplification of IntI1, dfrA5, dfrA7 and dfrA25 genes
Table (4): Molecular detection of IntI1, dfrA5, dfrA7 and dfrA25 genes
Genes |
Positive (%) |
Negative (%) |
P-value |
|---|---|---|---|
IntI1 |
62 (31%) |
138 (69%) |
0.0251 |
dfrA5 |
20 (10%) |
180 (90%) |
0.0023 |
dfrA7 |
34 (17%) |
166 (83%) |
0.0054 |
dfrA25 |
24 (12%) |
176 (88%) |
0.0031 |
IntI1-dfrA5 |
13 (7%) |
187 (93%) |
0.0019 |
IntI1-dfrA7 |
20 (10%) |
180 (90%) |
0.0023 |
IntI1-dfrA25 |
12 (6%) |
188 (94%) |
|
IntI1-dfrA5-dfrA7 |
14 (7.5%) |
191 (92.5%) |
|
IntI1-dfrA5-dfrA7-dfrA25 |
11 (5.5%) |
189 (94.5%) |
|
dfrA5-dfrA7 |
20 (10%) |
180 (90%) |
0.0023 |
dfrA5-dfrA7-dfrA25 |
13 (7%) |
187 (93%) |
0.0019 |
dfrA5-dfrA25 |
12 (6%) |
188 (94%) |
|
dfrA7-dfrA25 |
16 (8%) |
184 (92%) |
0.0055 |
Antibiotic resistance pattern analysis is summarized in Table 5. More than 60 different resistance profiles were identified, of which 37 were common patterns. Among isolates carrying the class 1 integron gene intI1, the most frequent resistance combinations were AMC-CEP (61 isolates), AMP-AMC-CEP (60 isolates), and AMP-AMC-TRI (60 isolates). Among isolates positive for dfrA5, the predominant patterns were AMP-AMC-TRI (14 isolates) and AMP-AMC-CEP (11 isolates). For dfrA7-positive isolates, AMC-TET and AMC-CEP were the most common patterns (17 and 16 isolates, respectively), whereas dfrA25-positive isolates were mainly associated with AMC-TET and AMP-AMC-CEP resistance profiles (13 and 12 isolates, respectively), as shown in Table 6.
Table (5): Distribution antibiotics resistance according to IntI1, dfrA5, dfrA7 and dfrA25 genes frequency
| Antibiotic agents | Resistance N (%) | Genes | P-value | |||
|---|---|---|---|---|---|---|
| IntI1 N (%) |
dfrA5 N (%) |
dfrA7 N (%) |
dfrA25 N (%) |
|||
| Chloramph-enicol | 72 | 9 (13%) | 1 (1.3%) | 3 (4.1) | 0 (0%) | 0.419 |
| Tetracycline | 180 | 15 (8.3%) | 7 (3.8) | 7 (3.8) | 9 (5%) | 0.551 |
| Lincomycin | 134 | 13 (9.7) | 3 (2.2%) | 9 (7%) | 11 (8.2%) | 0.472 |
| Ampicillin | 198 | 15 (8%) | 4 (2%) | 10 (5%) | 13 (7%) | 0.573 |
| Amoxicillin | 199 | 17 (9%) | 5 (3%) | 9 (5%) | 8 (4%) | 0.4444 |
| Cefotaxime | 186 | 6 (3.2) | 5 (3%) | 9 (5%) | 7 (4%) | 0.658 |
| Ceftriaxone | 98 | 3 (3%) | 0 (0%) | 4 (4%) | 2 (2%) | 0.675 |
| Cephalexin | 200 | 8 (4%) | 2 (1%) | 8 (4%) | 9 (5%) | 0.601 |
| Nalidixic acid | 108 | 7 (6.4) | 6 (6%) | 7 (6.4) | 5 (5%) | 0.892 |
| Trimethoprim | 102 | 14 (14%) | 10 (10%) | 11 (11%) | 13 (13%) | 0.772 |
| Ciprofloxacin | 52 | 4 (8%) | 0 (0%) | 0 (0%) | 2 (4%) | 0.104 |
| Amikacin | 43 | 3 (7%) | 1 (2.3) | 1 (2.3) | 6 (14%) | 0.096 |
| Gentamicin | 42 | 6 (14.2%) | 0 (0%) | 3 (7.1) | 2 (5%) | 0.065 |
| Imipenem | 34 | 2 (6%) | 1 (3%) | 1 (3%) | 3 (9%) | 0.494 |
Table (6): Antibiotics resistance patterns according to IntI1, dfrA5, dfrA7 and dfrA25 genes cassette array frequency
No. |
Resistance patterns |
IntI1 (n = 62) |
dfrA5 (n = 20) |
dfrA7 (n = 34) |
dfrA25 (n = 24) |
|---|---|---|---|---|---|
1. |
AMC-TET |
51 |
10 |
17 |
13 |
2. |
CEP-C |
47 |
3 |
6 |
1 |
3. |
LIN-TET |
45 |
4 |
7 |
7 |
4. |
AMP-AMC-LIN |
44 |
4 |
8 |
5 |
5. |
AMP-AMC-TRI |
60 |
14 |
9 |
11 |
6. |
AMC-CEP |
61 |
9 |
16 |
9 |
7. |
AMP-AMC-CEP |
60 |
11 |
12 |
12 |
8. |
AMC-LIN |
44 |
8 |
12 |
3 |
9. |
CEP-AMC-C |
32 |
9 |
6 |
5 |
10. |
CEP-LIN |
36 |
10 |
10 |
3 |
11. |
CEP-C |
47 |
6 |
12 |
9 |
12. |
LIN-C-IME |
29 |
2 |
0 |
1 |
13. |
AMP-AMC-CEP-C |
51 |
8 |
5 |
6 |
14. |
AMC-CEP-TET-AMP |
58 |
10 |
7 |
7 |
15. |
AMP-AMC-C-IME-TRI |
32 |
4 |
1 |
2 |
16. |
AMC-CEP-IME-AMP |
31 |
1 |
4 |
2 |
17. |
AMC-CEP-GEN-AMP |
33 |
8 |
2 |
5 |
18. |
LIN-AMC-CEP-AMP |
31 |
0 |
3 |
3 |
19. |
NAL-AMC-CEP-TET |
42 |
7 |
9 |
6 |
20. |
AMC-CEP-LIN-AMP,AMI |
37 |
5 |
1 |
4 |
21. |
CIPR-AMC-CEP-LIN-GEN |
40 |
0 |
0 |
2 |
22. |
AMC-CEP-LIN-C-TET |
44 |
9 |
10 |
7 |
23. |
AMC-CEP-LIN-GEN |
39 |
3 |
5 |
3 |
24. |
AMC-CEP-C-TET-CIP |
41 |
0 |
0 |
2 |
25. |
AMC-CEP-TET-GEN-TRI |
37 |
9 |
5 |
9 |
26. |
CIP-NAL-AMC-CEP-GEN |
28 |
4 |
7 |
3 |
27. |
NAL-AMC-CEP-LIN-C-TRI |
19 |
3 |
7 |
2 |
28. |
AMP-AMC-CEP-LIN-TET-TRI |
23 |
5 |
2 |
1 |
29. |
AMC-CEP-LIN-C-TET |
19 |
9 |
11 |
6 |
30. |
AMC-CEP-LIN-GEN-AMI |
23 |
6 |
3 |
2 |
31. |
AMP-AMC-CEP-LIN-C-TET-AME-TRI |
20 |
2 |
2 |
2 |
32. |
AMP-AMC-CEP-LIN-C-GEN |
17 |
5 |
5 |
2 |
33. |
AMP-AMC-CEP-C-TET-GEN |
22 |
7 |
3 |
4 |
34. |
CIP-NAL-AMP-AMC-CEP-C-TET-AME-CEX |
14 |
0 |
0 |
1 |
35. |
AMP-AMC-CEP-LIN-CLR-TET-GEN |
10 |
2 |
1 |
3 |
36. |
CIP-NAL-AMP-AMC-CEP-LIN-CLR-C-TRI, CEX-IMP |
9 |
0 |
0 |
2 |
37. |
CIP-NAL-AMP-AMC-CEP-LIN-IME-C-TET-TRI-CEF-IMP |
5 |
0 |
0 |
3 |
Multiple Drug Resistance (MDR) 94% |
98% |
84% |
97% |
89% |
*Abbreviations: Chloramphenicol (C), Tetracycline (TET), Lincomycin (LIN), Ampicillin (AMP), Amoxicillin (AMC), Cefotaxime (CEF), Ceftriaxone (CEX), Cephalexin (CEP), Nalidixic acid (NAL), Trimethoprim (TRI), Ciprofloxacin (CIP), Amikacin (AME), Gentamicin (GEN), Imipenem (IMP)
The global rise of multidrug-resistant K. pneumoniae has made treatment of nosocomial and community-acquired infections increasingly difficult.17 Mobile genetic elements, particularly integrons, play a major role in the acquisition and dissemination of antimicrobial resistance genes among Enterobacteriaceae.18 In the present study, most isolates showed high resistance to commonly used antibiotics such as cephalexin, ampicillin, amoxicillin, and cefotaxime, whereas lower resistance rates were observed for imipenem, gentamicin, amikacin, and ciprofloxacin. Similar findings have been reported in studies from Iran, Iraq, and India, although resistance rates vary depending on local antibiotic use and infection control practices.19-21 Misuse and over the counter availability of antibiotics in many regions, including Iraq, may contribute to the rapid selection and spread of resistant strains.
Molecular analysis showed that 31% of isolates carried the class 1 integron gene intI1, supporting the role of integrons in the dissemination of multidrug-resistance. This prevalence was lower than reports from northwest Iran and other regions, where higher integron frequencies have been observed, possibly reflecting differences in antimicrobial pressure, patient populations, and infection control practices.22 Nevertheless, integron positive isolates in our study tended to show higher resistance rates, consistent with previous reports demonstrating the association between integrons and MDR phenotypes.22
Among the resistance gene cassettes examined, dfrA7 was the most frequently detected, followed by dfrA25 and dfrA5. These genes encode trimethoprim resistant dihydrofolate reductase enzymes and are commonly found within class 1 integrons of Enterobacteriaceae.13 Similar cassette distributions have been reported in studies from China and Iran, although the predominant cassette types vary geographically.16,23-27 The higher prevalence of dfrA7 in our isolates may be related to selective pressure from trimethoprim sulfamethoxazole use or to the presence of highly transmissible mobile genetic elements carrying this cassette. Horizontal transfer of integron associated gene cassettes through plasmids and transposons likely contributes to the persistence and spread of these resistance determinants.
We also identified isolates harboring multiple gene cassettes (intI1-dfrA5-dfrA7-dfrA25), indicating accumulation of resistance determinants within single integrons. Such cassette arrays have been reported previously and are associated with increased multidrug-resistance.28,29 However, the presence of a resistance gene does not always correlate with phenotypic resistance, which may result from weak promoter activity, gene mutations, or regulatory mechanisms affecting gene expression.30
Overall, the high prevalence of multidrug-resistant K. pneumoniae and the detection of class 1 integrons in this study highlight the importance of continuous surveillance of resistance genes and rational antibiotic use. Strengthening antimicrobial stewardship programs and monitoring integron associated resistance determinants are essential strategies to control the spread of resistant uropathogens.30
This study demonstrated a high prevalence of multidrug-resistant K. pneumoniae among urinary tract infection isolates and confirmed the presence of class 1 integrons and associated resistance gene cassettes. Among the cassette genes examined, dfrA7 was the most frequently detected, highlighting the contribution of integron-mediated gene transfer to trimethoprim resistance and overall antimicrobial resistance in these isolates. The association between integron carriage and elevated resistance rates underscores the importance of monitoring mobile genetic elements in clinical pathogens. Continuous molecular surveillance of integrons and resistance gene cassettes, together with rational antibiotic use and strengthened antimicrobial stewardship, is essential to limit the spread of resistant K. pneumoniae. Further research into novel antimicrobial agents and alternative therapeutic strategies is needed to address the growing threat of antimicrobial resistance.
ACKNOWLEDGMENTS
The authors extend their thanks and gratitude to Al-Rifai Teaching Hospital in Dhi Qar province for facilitating the collection of samples. The authors also extend their thanks to everyone who contributed to the completion of this research.
CONFLICT OF INTEREST
The authors declare that there is no conflict of interest.
AUTHORS’ CONTRIBUTION
All authors listed have made a substantial, direct and intellectual contribution to the work, and approved it for publication.
FUNDING
None.
DATA AVAILABILITY
All datasets generated or analyzed during this study are included in the manuscript.
ETHICS STATEMENT
Not applicable.
INFORMED CONSENT
Written informed consent was obtained from the participants before enrolling in the study.
- Priyadarshini A, Kalola P, Patadia H, Shah J, Gangawane A. Prevalence, aetiology and antimicrobial resistance profile of diabetic individuals suffering from community-acquired urinary tract infection. J Med Microbiol. 2025;74(1).
Crossref - Khadhim MK, Kazaal MA. Antibiotics Resistance and Integron Class 1 among Commonsal Escherichia coli. AL-Qadisiya Medical Journal. 2017;13(24):1-10.
Crossref - Kharel S, Rimal S, Neupane B, et al. Detection of AcrAB efflux pump mediated ciprofloxacin resistance in Escherichia coli and Klebsiella pneumoniae in Nepal. BMC Infect Dis. 2025;25(1):1227.
Crossref - Kazaal MA, Hamad WA, Atiya WH, Saeed BJ, Abd-Alsatar AN. Impact of Antibiotic Resistance on Sustainable Development Goals. AIP Conf Proc. 2023;2776:020016.
Crossref - Olaitan MO, Orababa OQ, Shittu RB, et al. Extended-spectrum beta-lactam-resistant Klebsiella pneumoniae in sub-Saharan Africa: a systematic review and meta-analysis from a One Health perspective. BMC Infect Dis. 2025;25(1):843.
Crossref - Lu S, Sheng J, Ter-Stepanyan MM, Wang Y, Mkrtchyan HV. Genomic characterization of an ESBL-producing Klebsiella pneumoniae ST37 recovered from a hospitalized patient in Armenia. Microbiol Spectr. 2025;13(9):e0033225.
Crossref - Gómez-Manzano FJ, Barroso-Corroto E, Laredo-Aguilera JA, Molina-Madueño RM, Rodríguez-Cañamero S, Carmona-Torres JM. Urination Stimulation Techniques for Collecting Clean Urine Samples in Infants Under One Year: Systematic Review and Meta-Analysis. Acta Paediatr. 2025;114(8):1788-1802.
Crossref - Jia L, Zhang G, Li X, et al. ESBL-producing Klebsiella pneumoniae isolated from dairy cows with pneumonia: insight into their epidemiology, genetic features and antimicrobial resistance profiles. Microb Pathog. 2025;207:107909.
Crossref - Namikawa H, Oinuma KI, Kaneko Y, Kakeya H, Shuto T. Antimicrobial resistance in hypermucoviscous and non-hypermucoviscous Klebsiella pneumoniae: a systematic review and meta-analysis. Emerg Microbes Infect. 2025;14(1):2438657.
Crossref - Cox PB, Teo JQ-M, Fouts DE, et al. Molecular epidemiology and clinical characteristics of carbapenem-resistant Klebsiella pneumoniae bloodstream and pneumonia isolates. Microbiol Spectr. 2025;13(8):e0063125.
Crossref - Yang X, Lou Z, Wang X, et al. Resistance profile and influence factors of carbapenem-resistant Klebsiella pneumoniae (CRKP) causing infections in China: a systematic review and meta-analysis. Ann Clin Microbiol Antimicrob. 2025;24(1):56.
Crossref - Qi S, Li H, Ma J, et al. Molecular characterization of colistin resistance in carbapenem-resistant Klebsiella pneumoniae from a tertiary hospital in China. Microbiol Spectr. 2025;13(10):e0104925.
Crossref - Jian Z, Liu Y, Wang Z, Liu P, Wang J, Yan Q, Liu W. Prevalence and molecular characteristics of colistin-resistant isolates among carbapenem-resistant Klebsiella pneumoniae in Central South China: a multicenter study. Ann Clin Microbiol Antimicrob. 2025;24(1):1.
Crossref - Chen T, Wang X, Xiong L, Shen P, Xiao Y. Emergence and molecular evolution of carbapenem-resistant hypervirulent ST23 Klebsiella pneumoniae: The superbug phenomenon in China. Virulence. 2025;16(1):2545556.
Crossref - Wang Q, Ye MY, Hong C, Li ZP, Lin L. The mechanisms of resistance, epidemiological characteristics, and molecular evolution of carbapenem-resistant hypervirulent Klebsiella pneumoniae. Lab Med. 2025;56(4):323-335.
Crossref - Firoozeh F, Mahluji Z, Khorshidi A, Mohammad Z. Molecular characterization of class 1, 2 and 3 integrons in clinical multi-drug resistant Klebsiella pneumoniae isolates. Antimicrob Resist Infect Control. 2019;8:59.
Crossref - Russo A, Fusco P, Morrone HL, Trecarichi EM, Torti C. New advances in management and treatment of multidrug-resistant Klebsiella pneumoniae. Expert Rev Anti Infect Ther. 2023;21(1):41-55.
Crossref - Wang L, Zhu M, Yan C, et al. Class 1 integrons and multiple mobile genetic elements in clinical isolates of the Klebsiella pneumoniae complex from a tertiary hospital in eastern China. Front Microbiol. 2023;14:985102.
Crossref - Jasim JS, Hussein AR. Detection of class 1 integron among Klebsiella pneumoniae clinical isolates in Baghdad hospitals. J Fac Med Baghdad. 2025;67(3).
- Al-Assil B, Mahfoud M, Hamzeh AR. First report on class 1 integrons and Trimethoprim-resistance genes from dfrA group in uropathogenic E. coli (UPEC) from the Aleppo area in Syria. Mob Genet Elements. 2013;3(3):e25204.
Crossref - Gidla VVS, Kumar VJA, Bhanu RV, Dinesh G. Integrons: a mobile genetic element of concern in antimicrobial resistance gene transfer: an overview. Indian J Vet Sci Biotechnol. 2023;19(3):1-5.
Crossref - Afzali H, Firoozeh F, Amiri A, Moniri R, Zibaei M. Characterization of CTX-M-Type Extend-Spectrum β-Lactamase Producing Klebsiella spp. in Kashan, Iran. Jundishapur J Microbiol. 2015;8(10):e27967.
Crossref - Rezaee MA, Langarizadeh N, Aghazadeh M. First report of class 1 and class 2 integrons in multidrug-resistant Klebsiella pneumoniae isolates from northwest Iran. Jpn J Infect Dis. 2012;65(3):256-259.
Crossref - Lina TT, Rahman SR, Gomes DJ. Multiple-antibiotic resistance mediated by plasmids and integrons in uropa- thogenic Escherichia coli and Klebsiella pneumoniae. Banglad J Microbiol. 2007;24(1):19-23.
Crossref - Li B, Hu Y, Wang Q, et al. Struc- tural diversity of class 1 integrons and their as-sociated gene cassettes in Klebsiella pneumoniae isolates from a hospital in China. PloS One. 2013;8(9):e75805.
Crossref - Omar F. H., A. H. Ibrahim. The Prevalence of Integron Class I and II among Multi-drug Resistance Producing Klebsiella pneumonia. Iraqi J. Agric. Sci. 2023;54(3):619-629
- Tekele SG, Mulatie Z, Gedefie A, et al. Prevalence of AmpC beta-lactamase producing Escherichia coli and Klebsiella pneumoniae in Africa: a systematic review and meta-analysis. Antimicrob Resist Infect Control. 2025;14(1):109.
Crossref - Salimizand H, Shahcheraghi F, Kalantar E, Badmasti F. Molecular characterization of class 1 in- tegrons and gene cassettes in multidrug resistant (MDR Klebsiella spp. Isolated from hospitalized and outpatients in Iran, 2009. Iran J Microbiol. 2013;5(1):48-55.
- Najjuka CF, Kateete DP, Kajumbula HM, Joloba ML, Essack SY. Antimicrobial susceptibility profiles of Escherichia coli and Klebsiella pneumoniae isolated from outpatients in urban and rural districts of Uganda. BMC Res Notes. 2016;9(1):235.
Crossref - Asghari B, Goodarzi R, Mohammadi M, Nouri F, Taheri M. Detection of mobile genetic elements in multidrug-resistant Klebsiella pneumoniae isolated from different infection sites in Hamadan, West of Iran. BMC Res Notes. 2021;14(1): 330.
Crossref
© The Author(s) 2026. Open Access. This article is distributed under the terms of the Creative Commons Attribution 4.0 International License which permits unrestricted use, sharing, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.