


Arbuscular mycorrhizal (AM) symbiosis is an important biological breakthrough which assisted plant land colonization over 400 million years ago. This widespread mutualistic interaction between fungi and plants enhances nutrient exchange, ecological sustainability, plant stress resistance, and host plant development. AM symbiosis improves plant nutrition by deriving nutrients through both mycorrhizal pathways and the Plant’s own pathways. AMF influence nutrient availability by altering soil properties, microbial populations, and nutrient cycling. Understanding the life cycle of AMF, spore germination, sporulation, colonization, and symbiosis formation are critical for large-scale agricultural applications. Root organ culture (ROC) techniques offer intriguing possibilities to mass producing AMF under in vitro. This review surveys the literature on these topics, focusing on methods for enhancing sporulation in in vitro. Enhancing in vitro sporulation can be achieved by supplementing growth media with phenolic compounds, fatty acids, and phytohormones, and optimizing the media and related factors. These compounds regulate fungal growth and development, leading to increased sporulation and improved AMF inoculant efficacy. Further research is needed to provide quality inoculum and develop crop-specific formulations and delivery methods to harness the potential of AMF in diverse agroecosystems.
AM Fungi, Life Cycle, Root Organ Culture, Signalling Compounds, Plant Hormones, Sporulation
Arbuscular mycorrhizal fungi form distinctive symbiotic partnerships with plants, with about seventy to ninety percent of plant species engaged in these mycorrhizal symbioses.1 For almost 400 million years, vascular plants and arbuscular mycorrhizal (AM) endophytes have coevolved to form close partnerships.2,3 Since they promote plant development and survival, these microorganisms are crucial to the ecology of the majority of vascular plants. Plants can contribute 4-20% of their photosynthate to support AM fungi, which translates to the consumption of approximately five billion tonnes of carbon per year by AM fungi.1,4 Host plants support the viability and effectiveness of mutualistic arbuscular mycorrhizal (AM) fungal partners by limiting sugar export while ensuring a supply of lipids to fungi. This strategy reduces the carbon costs linked to parasitism while still benefiting from the nutrient uptake facilitated by AM fungi.5 Therefore, it is likely that specific molecules, such as fatty acids, rather than the total carbon flow, influence the carbon-for-phosphorus exchange between AM fungus and the host plant.
Mutualistic relationships between plants and fungi result in colonizing the root cortex, allowing for the exchange of minerals, especially phosphorus, and carbon between the fungus and the plant. For a long time, the large-scale production of contaminant-free AM fungi has presented an obstacle to their application in agriculture.
Owing to the biotrophic nature of AM fungi, the challenges connected with inoculum production have severely limited their utilization in agriculture. The most common method for AM production involves growing host plants with AM inoculation and in turn chopping mycorrhizal roots and mixing them with a growing medium that contains hyphae and spores. Lesser spore production and contaminants are the major quality issues associated with this type of inoculum. Additional techniques have been developed in an effort to improve quality and dependability, such as aeroponic culture6,7 and alginate encapsulation of root pieces.8 The most promising method for producing bulk quantities of pathogen-free, high-quality AM inoculum, which is desired for application in various crop cultivation is still propagule generation under aseptic conditions.
However, the monoxenic cultivation approach has evolved into an effective technique for generating contaminant-free AM fungi, facilitating large-scale production under rigorously controlled conditions, starting from the initial research of Mosse and Hepper9 and later advancements by Strullu and Romand,10,11 as well as Becard and Fortin.12 Innovative fundamental research has been carried out with various AM fungal species that have been effectively cultured from root organs.13 With respect to inoculum production, monoxenic cultivation offers a number of advantages over conventional cultivation methods. This method provides contaminant-free, clean, sterile, bulk propagules that were previously impractical for production through conventional/ hydroponic/aeroponic methods. Furthermore, this technology facilitates the production of many spores within a shorter period of time with less space. Although various methods are being used for inoculum production the common issue is spore production. Spores are considered essential for developing subsequent generations and follow various events for the process of sporulation. Many options exist to increase the potential for AM fungal sporulation and propagule development via both methods of production. This review provides a literature survey and concise details regarding AM fungi life cycle, in vitro production methods, constraints and suggestions to overcome and approaches for enhancing sporulation and propagule development, which are essential for large scale AM production.
Arbuscular mycorrhizal fungal life cycle and symbiosis establishment
AM fungi have a multistep life cycle that depends on plants and requires fungus-host root recognition, signalling, and communication with one another.
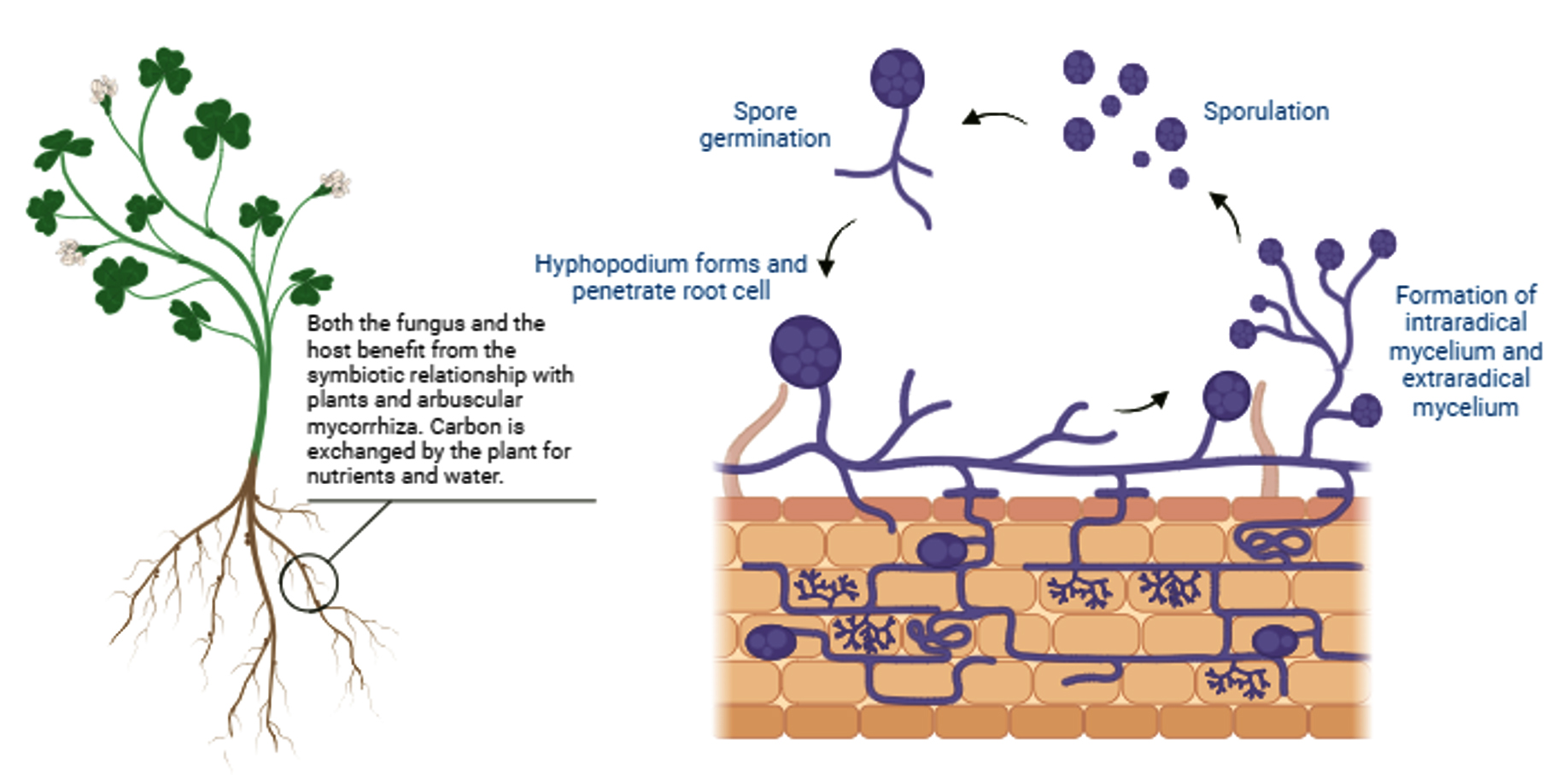
Figure 1. Life cycle of arbuscular mycorrhizal fungus. The graphic depicts critical processes such as spore germination, hyphal development, root colonization, and the emergence of arbuscules and vesicles inside plant roots. The symbiotic interaction between AMF and plant roots improves nutrient uptake and plant resistance. Both the fungus and the host benefit from the plants’ symbiotic relationship with arbuscular mycorrhiza. Carbon is exchanged by the plant for nutrients and water
Figure 1 depicts the processes of root colonization by AM fungus. The life cycle of AM fungus begins with the spontaneous germination of fungal spores in soil under favourable environmental circumstances, in the absence of a host plant.14,15 Fungal colonies grow several centimetres and develop distinct growth structures.16 This asymbiotic phase transforms into a presymbiotic phase, which is characterised by considerable hyphal branching caused by the presence of the host plant.17 During the presymbiotic phase, strigolactone, a plant hormone secreted from roots,18 attracts AM fungus. Strigolactones activate the metabolism of AMF and stimulate spore germination and hyphal branching to facilitate root colonization.19 In response to plant signals, AMF produce LCOs and COs, called Myc factors. When AM fungal mycelia directly touch the root cortex’s cell surface and form an appressorium, presymbiosis occurs.20 Appressorium formation signifies the start beginning of the fungus-host plant symbiotic relationship. The creation of internal arbuscules creates a large surface area for nutrient exchange between fungal-acquired soil nutrients and the host, which is required to initiate the interaction.21 When the hyphae penetrate the cell walls and develop into tree-like structures known as arbuscules within the cortical cells by recurrent dichotomous branching, the plant plasma membrane converts into the peri-arbuscular membrane, which serves as the site of nutrition exchange.22 The hyphae multiply in the soil, develop extraradical mycelia, and complete their life cycle by producing new asexual spores in extraradical mycelia.23 Extraradical mycelia extend and generate additional spores for later germination by responding to exudate signals and completing their life cycle.
AM fungal inoculum production
AM fungi are increasingly being used in horticulture, agriculture, and forestry, as well as to remediate the environment, improve crop health and productivity, and reduce pesticide use.24,25 Nonetheless, the development of large-scale production strategies to acquire high-quality AM fungal inoculum has been limited by the mandatory biotrophic nature of this fungus, which also limits its commercial use.
Over the last few decades, numerous technologies and products have been produced, each with its own set of advantages and disadvantages in terms of design and application. AM fungus are produced in huge quantities using three well-known techniques: (1) Substrate-based production systems are common and principally offer an economical way of manufacturing AM fungal inoculum in substantial quantities for use in large-scale applications. (2) Substrate-free production technology designed to generate a relatively contaminant-free (sheared) AM fungal inoculum. This can be accomplished through aeroponic or hydroponic methods. However, the higher costs of these manufacturing processes have generally restricted their employment to research and small-scale applications. (3) In vitro production strategies rely on either excised roots or root organ cultures. Although these technologies are expensive, they ensure the production of more infective units and contaminant-free AM fungal inoculum.26 Figure 2 depicts the various ways for bulk production of AM fungus.
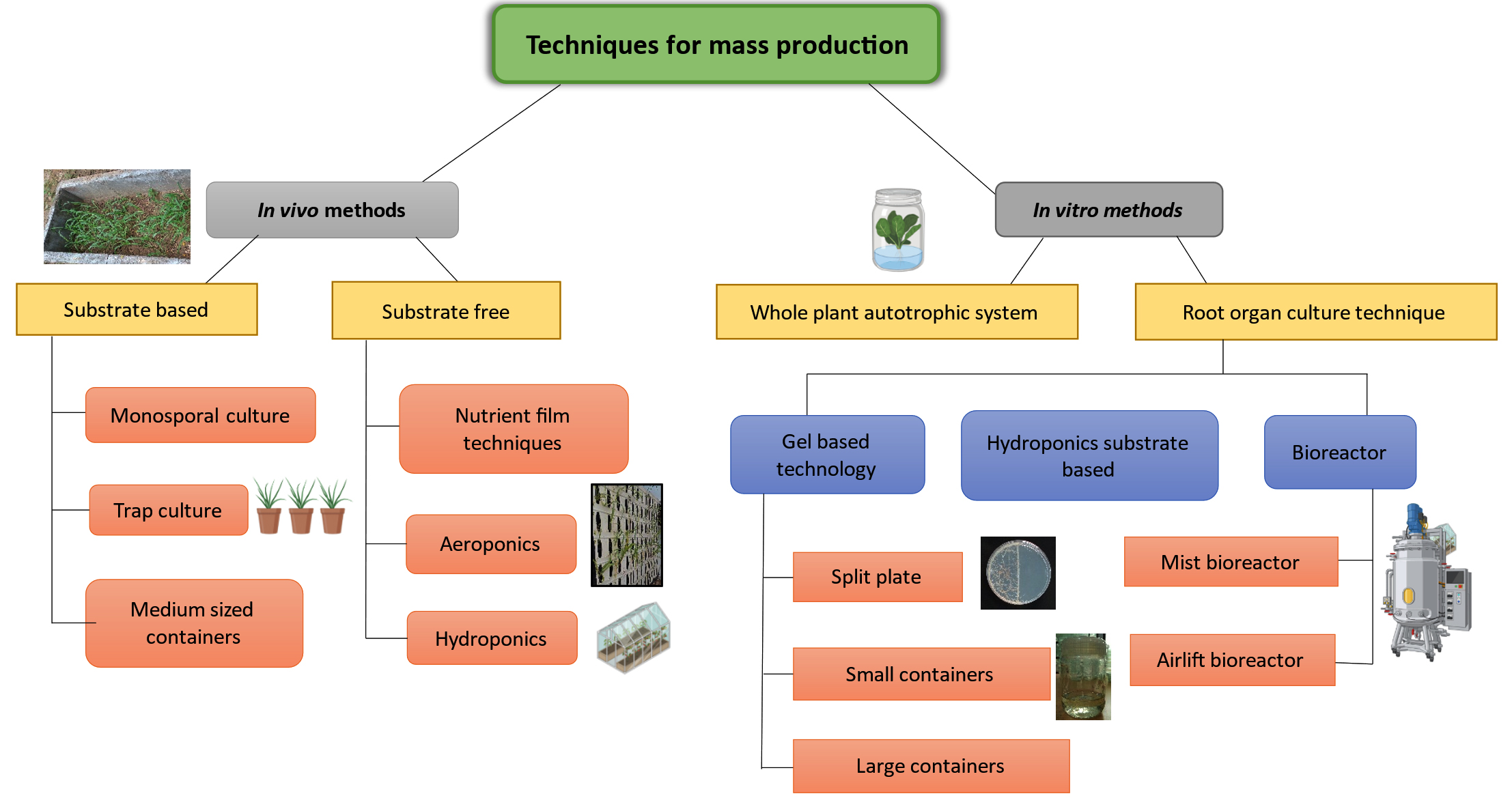
Figure 2. Techniques for the mass production of AM fungi. It includes growing host plants in sterilized soil (pot cultures), using root organ culture systems in controlled lab conditions (in vitro cultures), and cultivating plants in air/mist environments (aeroponic systems) to enhance root colonization. Additionally, scalable production can be achieved through bioreactors utilizing liquid culture systems or solid-state fermentation. Specialized soil conditions are also maintained in field or greenhouse systems for effective mass production of AM fungi
Conventional methods of AM production
Conventional ways for mass multiplying AM fungal inoculum include substrate-based production systems (nursery beds, pots, concrete tanks, etc.) and substrate-free production systems (aeroponics).27
Substrate based production method
Conventional substrate-based inoculum production is the most popular traditional technique for large-scale production of AM fungus. Commonly used substrates for production include vermiculite, perlite and zeolite. However, these methods have serious issues with cross-contamination during the process; therefore, producing high-quality inoculum continues to be a challenging endeavour.28
The most prevalent approach for producing AM fungal inoculum is substrate-based production of AM fungi in beds, bags, or pots which requires minimal technical support and uses affordable consumables.29 The media that are homogeneous in composition, aid in aeration and retain enough water for plant growth are vermiculite, calcinated clay, perlite and expanded clay which are considered for AM inoculum production. Even with stringent control measures in place, substrate-based growing systems are rarely able to guarantee the absence of contaminants. Furthermore, these methods usually demand considerable space and pest management.26
A single species or a group of identified AM fungal species in clay or plastic pots, or scaled up to medium-size bags and containers, and large raised or grounded beds, are typically used to begin mass production in sand/soil or other substrate-based production methods.30,31 On-farm production is sometimes started using species that are native to the application site and are not always identified to the species level. With regard to soil aeration, drainage, oxygen delivery, and root growth, Gaur and Adholeya produced the most AMF inoculum in sand,32 which may have allowed for superior soil aeration. The low maintenance cost of growing on sand is another advantage.33 This kind of technology allows for the appropriate monitoring and control of the nutrient delivery to AM fungi and plants. Though there may be a chance of excessive contaminants, this method might offer regulated culture conditions.28
Substrate free production
A few techniques, such as the nutrient flow technique, hydroponics, and aeroponics systems, are known to produce AM inoculum without the use of substrates. Regarding how the nutritional solution is applied and how it is aerated, all of these methods may vary from one another. The roots of the host plant are submerged in a solution of water and nutrients when growing hydroponically. To keep the roots from experiencing a shortage of oxygen, an aeration pump oxygenates the nutritional solution in this static type of system. To reduce the flow of nutritional solutions and the buildup of air bubbles, which could harm the development of fragile extraradical hyphae, the pumps must be turned on sometimes.26,34 Mosse and Thompson introduced the nutrient flow approach,35 which Lee and George later reviewed.36 By covering the roots with a thin layer of nutritional solution, this method provides an alternate strategy that increases the surface area available for gas exchange and solves problems associated with insufficient aeration in the sloped channels where arbuscular mycorrhizal (AM) fungus and plant roots develop.
Aeroponic cultivation is a technique for growing plants without the use of soil, where the roots hang in the air and are nourished by a nutrient-rich mist. This differs from hydroponics, which involves submerging plant roots in a solution of water and nutrients. Zobel et al. first used aeroponic culture to study the relationship between rhizobia and legumes,37 and subsequently for Sylvia and Hubbell’s research on VAM fungi.7 To culture VAM fungi in an aeroponic environment, seedlings that have already been inoculated and colonized are moved to the aeroponic setup to promote fungal colonization, extensive root growth, and sporulation. In this method, plants are grown via a fine spray of nutrient solution that helps maintain moisture in the roots, promoting colonization. Compared with conventional hydroponics, this system offers a more oxygen-rich environment compared to conventional hydroponics and has been shown to effectively produce VAM inoculum without the need for a physical substrate.6
In vitro culture methods for AM production and enhanced sporulation
Root organ culture of AMF
Root organ culture refers to a technique for cultivating sterile, complete plant roots, excluding shoots, in nutrient-rich media. This technique is valuable for investigating AM fungal colonization and growth characteristics. The association between an endogone species and a plant was originally reported by Mosse, not long after the first attempts to culture AM fungus in vitro,38 which began in the late 1950s.39 Arbuscular mycorrhizal (AM) fungi were effectively grown from excised tomato (Lycopersicum esculentum Mill) roots on gelled media by Mosse and Hepper in the middle of the 1970s.9 By using Ri T-DNA-transformed carrot roots as hosts, Mugnier and Mosse and, Becard and Fortin introduced “ROC systems” ten years later, increasing the spore production of AM fungi.12,40
When the soil bacteria A. rhizogenes infects plant tissues, it can cause a neoplastic disease or adventitious roots, sometimes known as hairy roots. The hairy root phenotype is caused by infection in plants when the T-DNA region from the root-inducing (Ri) plasmid of A. rhizogenes is inserted into the DNA of the plant. The T-DNA portion of the Ri-plasmid is integrated into the nuclear genomes of hairy roots.41 These hairy roots exhibit distinct characteristics, including frequent branching, a high density of capillary roots, substantial biomass production capacity, and rapid growth. Their susceptibility to infection by A. tumefaciens, A. rhizogenes, and other bacteria that can transfer genes to plants varies significantly throughout plant species.42 Roots from Convolvulus sepium and Daucus carota that were transformed by Tepfer and Tempe using Ri-T DNA have been used for AM culture.43
The ROC of AM isolates solve the problem of producing large amounts of inoculum. It provides a pure, viable, and contamination-free inoculum while taking up less space, giving it an advantage over conventional pot-culture multiplication techniques. Using various host plants and AM fungal inoculants, Table 1 shows how AM fungi can be grown in a monoxenic environment.
Table (1):
AM fungi cultured in the monoxenic system with different host plants and AM fungi
Host plant |
Fungal inoculum used |
Ref. |
|---|---|---|
Tomato (Lycopersicum esculentum Mill.) Red clover (Trifolium pratense L.) NT |
Glomus mosseae (Nicolson & Gerdemann) |
9 |
Carrot (Daucus carota L.) Bindweed (Convolvulus sepium L.) T |
Glomus mosseae |
40 |
Solanum lycipersicum L. NTDaucus carota L. T |
Glomus versiforme (Karsten) Berch |
44,45 |
Solanum lycopersicum L. NTDaucus carota L. T |
Glomus intraradices (Schenck & Smith) |
11,46 |
Daucus carota L. T |
Glomus macrocarpum (Tulasne & Tulasne) |
45 |
Daucus carota L. T |
Glomus etunicatum (Becker & Gerd) |
47 |
Sugarcane (Saccharum officinarum) T Sorghum (Sorghum vulgare) T |
Glomus mosseae (Nicolson & Gerdemann) |
48 |
Daucus carota L. T |
Glomus intraradices |
49 |
Carrot (Daucus carotaL.) T |
Glomus irregulare (DAOM-197198G. cerebriforme (DAOM-227022), G. aggregatum (strain 2101-sp) and G. diaphanum (DAOM-229456) from the Glomeromycota In vitro Collection (GINCO). |
50 |
Cassava (Manihot esculenta Crantz) T |
Rhizophagus irregularis |
51 |
Carrot (Daucus carota L.) T |
Glomus intraradices |
52 |
Tomato (Solanum lycopersicum L.) T |
Gigaspora gigantea (T.H. Nicolson & Gerd) |
53 |
Chicory (Cichorium intybus L.) T |
Strain INCAM 11, DAOM 711363 Rhizophagus irregularis from (INCA) |
54 |
Carrot (Daucus carota L.) T |
Rhizophagus species (SSDP005). |
55 |
Tomato (Solanum lycopersicum L.) T |
Rhizophagus irregularis (00101SP, MUCL43194) from SYMPLANTA (Germany) |
56 |
Daucus carota L. T |
Rhizophagus irregularis (DAOM 197198) |
57 |
Daucus carota L. T |
R. irregularis Schenk and Smith (DAOM, 197198; Biosystematics Research Center, Ottawa, Canada) |
58 |
Daucus carota L. T |
Rhizophagus clarus, Rhizophagus irregularis Rhizophagus aggregatus MUCL49408, Rhizophagus intraradices MUCL 49410 and Glomus hoi (Berch and Trappe) MUCL 45686 were provided by the Glomeromycota in vitro collection (GINCO, Belgium) |
59 |
Alkanna tinctoria |
Rhizophagus irregularis MUCL 41833 and MUCL 43194 (GINCO) |
60 |
Crucially, the absence of normal photosynthetic components and normal hormonal interactions results in a genetically modified root structure that replaces the natural plant host. It’s possible that genetically altered root structures won’t survive the long number of subcultures required for continuous production. Sucrose is added to the culture medium to make up for the absence of photosynthates, which may change the way the plant and fungal interact biochemically.13 The generation of hairy roots and AM colonisation are directly impacted by the sugar concentration, making it essential. These sugar concentrations may have an impact on the development of arbuscules, vesicles, and spore germination in in vitro converted carrot roots.
An efficient method for producing different AMF isolates on a wide scale under ROC circumstances is coculturing and it leads to the generation of substantial inoculum. When implemented in the rhizosphere of plants, it can offer multiple advantages due to its more diverse mycorrhizal population, which is typically found at relatively low densities in tropical soils. The possibility of large-scale production is made possible by the first successful cocultivation of the two genera, Glomus intraradices (Smith and Schenck) and Gigaspora margarita (Becker and Hall), within carrot roots.61 The main differences between AM spore production in vivo and in vitro are outlined in Table 2, with respect to their respective habitats, dependencies, controllability, scale, and applications.
Table (2):
Distinctions between in vivo and in vitro AM inoculum production in terms of their environments, dependencies, controllability, scale, and applications
Aspects |
In vivo spore production |
In vitro spore production |
|---|---|---|
Definition |
Spore production occurs naturally within the host plant’s rhizosphere, typically in soil as well as with the use of substrate |
Spore production is induced and controlled in laboratory conditions outside of the host plant. |
Environment |
Takes place within the natural soil environment surrounding the host plant. |
Occurs in controlled laboratory conditions, often on artificial growth media. |
Host plant dependency |
Dependent on the presence and health of the host plant. |
Spore production can be initiated without the presence of a host plant or with a limited host association. |
Regulation and manipulation |
Less controllable and subject to environmental factors. |
More controllable and can be manipulated through nutrient levels, pH and other growth conditions. |
Scale |
Scale is limited to the natural conditions of the plant’s environment. |
Production can be scaled up or down based on laboratory capacity and requirements. |
Applications |
Commonly used for largescale production |
Utilized in laboratory research, biotechnology, and commercial production of AM fungi. |
Factors influencing AM development under ROC
Host root selection
Both spore generation and fungal colonization depend heavily on host explants. Variable hosts both within and between genera have a major impact on mycorrhization occurrences. The influence of the host on mycorrhizal colonization has been widely studied under in vivo conditions for inoculum production. The choice of suitable Ri T-DNA transformed host roots enhances the mass production technique for monoxenic culture. Very few studies have been conducted under in vitro conditions.
Tiwari and Adholeya investigated the symbiotic interactions of AM fungi in the presence of hosts and analysed the nutritional dynamics of AM-host pairings under in vitro conditions. This study utilized four distinct carrot cultivars and clover (Trifolium subterraneum) transformed root cultures, along with the inoculation of Glomus intraradices. The findings revealed differences in mycorrhizal characteristics, such as sporulation, vesicle production, and both intraradical and extramatrical spread of the fungus, highlighting the influence of the host on the varying expression of G. intraradices, which accounts for the selective specificity of AM fungi toward hosts in vitro. Differences in the mechanisms of mycorrhizal spread and sporulation were noted even among various cultivars of the same host.62
Root systems exhibiting dense colonization and supporting extensive extramatrical mycorrhizal networks could be considered optimal candidates for large-scale inoculum production. These studies may play crucial roles in creating industrial-scale AM inocula for field use, utilizing suitable combinations of AM and host species. The identity of the host significantly influences the performance of various AM fungi.63,64 For example, the composition of nucleotypes in spores from heterogeneous R. irregularis strains varies on the basis of the host.65 The symbiotic relationship can lead to differing transcriptional responses from each partner, influenced by the genotypes of both the host and fungus.66 Moreover, the introduction of a different bacterial strain into the same host cultivar can alter its susceptibility to mycorrhizae.67 Given the close relationship between host identity and fungal performance, the successful culture of AM species in a monoxenic system requires an evaluation of how increased diversity of AM species impacts root variation in hosts.
Root systems exhibiting dense colonization and supporting extensive extramatrical mycorrhizal networks could be considered optimal candidates for large-scale inoculum production. These studies may play crucial roles in creating industrial-scale AM inocula for field use, utilizing suitable combinations of AM and host species. The identity of the host significantly influences the performance of various AM fungi.63,64 For example, the composition of nucleotypes in spores from heterogeneous R. irregularis strains varies on the basis of the host.65 The symbiotic relationship can lead to differing transcriptional responses from each partner, influenced by the genotypes of both the host and fungus.66 Moreover, the introduction of a different bacterial strain into the same host cultivar can alter its susceptibility to mycorrhizae.67 Given the close relationship between host identity and fungal performance, the successful culture of AM species in a monoxenic system requires an evaluation of how increased diversity of AM species impacts root variation in hosts.
Type of growth media
The type of growth medium utilized has a crucial effect on the production of AM propagules in vitro. The induction and growth of adventitious root cultures are impacted differently by different culture media.68 In mass production systems that aim for high propagule recovery, the growth medium becomes even more critical. Adjusting the medium can greatly increase the output of in vitro systems69; however, before pursuing commercial production, it is crucial to take the system’s feasibility and cost-effectiveness into account.
Minimal (M) medium has been extensively utilized for in vitro studies of AM fungi; however, it seems inadequate for cultivating all species of AM fungi.70 Adjusting the composition and pH of the medium for specific fungal isolates could enhance our understanding of the intricate biology involved in this symbiotic relationship.
The modified White’s medium (M medium)12 and the modified Strullu and Romand medium (MSR medium)71 are regarded as the two most effective and commonly used media for in vitro mycorrhizal root cultures.72 The M medium was initially created for tomato root organ cultures and represents a modification of White’s medium.73 Compared with Murashige and Skoog media (MS) and B5 media, White’s media feature a considerably lower microelement composition and are often employed for in vitro plant development.74 The M medium was developed based on a bioassay that evaluated the effects of different element concentrations on mycorrhiza synthesis, even though this diluted medium is sufficient for root development.12 A modified version of the MSR medium was developed to promote the growth of the fungus’s intraradical stage in in vitro.45 In terms of macroelement composition, the MSR and M media are similar. However, the concentrations of oligo elements and vitamins vary between the two: the MSR medium does not contain iodine, myo-inositol, or glycine, whereas the M medium lacks pantothenate, biotin, and cyanocobalamin. Since the absence of these components in either medium does not visibly affect AM symbiosis, their presence may not be strictly essential.75
The type of media utilized, as well as the concentration of carbon sources, influences root growth in root organ culture. Under laboratory conditions aimed at increasing plant photosynthesis for carbon synthesis, several carbon sources are added to root organ cultures, with sucrose being demonstrated to strongly stimulate the growth of adventitious root biomass.76 MSR media without sugar increased the in vitro germination rates for R. irregularis.77 Furthermore, In vitro AM spores have already been produced in bioreactors using liquid M media,78 and in vitro growth processes require adequate aeration of the liquid medium. The semisolid media exhibited the highest rates of AM root colonization and root development, with the porous and liquid media coming in second and third. In S. lycopersicum roots grown in in vitro, Ellatif et al. found that the number of arbuscules and the rate of mycorrhizal colonization were higher on solid media than on liquid media.53
pH of the growth media
Numerous organic buffers have been added to nutrient solutions in media for monoxenic cultures because of the significance of pH in the development of mycorrhizal symbioses.79 According to Vilarino et al. buffers promote the growth of arbuscular mycorrhizal fungus (AMF) external hyphae, which promotes sporulation.80 Douds modified the buffer’s pH and the medium’s phosphorus levels to enhance the mycorrhizae’s successful development using carrot plants and Funiliformis mosseae in in vitro. They observed that, despite the hyphae spreading throughout the Petri dishes, cultures kept for more than 17-24 weeks did not generate new spores.70 Additionally, Pawlowska et al. observed that F. mosseae did not sporulate during dual cultures. According to one study, the ideal pH range for AM fungal sporulation is between 6.0 and 7.0.47 In particular, AM fungus consistently generates spores most efficiently at pH 6.0, although sporulation is reduced at both lower and higher pH values.81 AM fungi thrive in conditions with pH values ranging from slightly acidic to neutral. According to research, the optimal pH range for the formation of AM fungal structures is between 5.5 and 7.0. The transformed root cultures (TRCs) used to propagate AM fungi, like Glomus intraradices and Gigaspora margarita, exhibit notable hyphal development and sporulation within this pH range. This highlights the importance of maintaining an optimal pH to maximize the effectiveness of AM fungal proliferation in vitro.61
Incubation conditions
In the root organ culture of AM fungus, light and dark incubation are also important. Differential light effects on AM-associated root colonization have been reported. Glomus claroideum colonized in vitro roots more when the environment was dark than when it was light, according to a study by Sahoo and Kullu.82 In contrast, the rate and percentage of AM infection in Allium cepa induced by Gigaspora calospora were lower at high light intensities (15-20 Klux) than at low light intensities (5-10 Klux).83 Similarly, it has been reported that larger AM arbuscules are produced at high light intensities than at low light intensities and that higher levels of photosynthetic photons are involved in the stimulation of root colonization by Glomus intraradices in Solanum tuberosum plantlets cultivated in an in vitro tripartite system.84 Although different reports exist, most of the studies used dark incubation
Dual culturing in bicompartments
It is possible to cultivate R. irregularis on dual-compartment plates in order to generate plant-free spores in in vitro.85 The fungal compartment (Fc) of a Petri plate can yield up to 34,000 spores using this culture method. Additionally, the interaction between the host and AMF determines the sporulation pattern in the hyphal compartment.86 Bicompartments allow for the exchange of signalling molecules and some nutrients, but they limit the spread of roots and mycelia between compartments. Exudates, such as sugars, amino acids, and other signalling chemicals, are released by the roots and diffuse into the fungal compartment via the barrier. These exudates are essential for the growth and development of AM fungus because they provide appropriate chemical signals and nutrients. In response to root exudates, fungal hyphae multiply and spread towards the source of the exudates (the root compartment), while remaining physically limited to their allocated compartment.44
A design was developed to enhance R. irregularis spore output in a dual-compartment culture system free from microbial and plant material contamination and to enable rapid production without requiring root snipping. The PVDF and cellophane culture techniques yielded a maximum of 61,395 and 49,155 spores per plate, respectively. Furthermore, these two culture methods can produce a greater number of spores than the usual culture system can produce, at a lower material cost and with much less labour. Low expenses and labour minimization are critical characteristics for an efficient cultural system. In the short-term, the cellophane culture method has emerged as the most inexpensive option.87 Alternative modifications to the traditional culture systems have been suggested, potentially leading to an average yield of 65,000 spores per plate. However, this method requires manual root trimming, a cultivation period of nine months, and, at times, complete media exchange in the fungal compartment every two months. In contrast, the two modified culture systems we examined produced nearly half the average number of spores per plate in just half the time.69 These spores can be utilized for developing inocula.
Breaking spore dormancy
For the production of spore inoculum, a limitation that arises with certain arbuscular mycorrhizal (AM) spore types is their dormancy. Dormancy in AM spores can lead to unsuccessful germination and hinder the development of fungal structures. Dormant spores do not germinate even when subjected to both physical and chemical conditions, which typically encourage germination and hyphal growth. Researchers have addressed the issue of spore dormancy in various studies. They applied cold stratification to break dormancy in AM spores during in vitro multiplication. Two primary factors contributing to this phenomenon were identified: the consistent age of the spores and the intrinsic dormancy present in newly produced spores. While the importance of cold stratification for overcoming natural dormancy is widely recognized, as noted by Mosse, applying cold treatment for over 14 days resulted in full germination with strong apical growth and minimal branching.88 Cold treatment of spores at 4 °C for 30 days increased germination rates to 56.5% and 90% in Glomus caledonium and Glomus intraradices, respectively.89 After establishing the symbiotic cultures, the extraradical fungal biomass began to develop and sporulate simultaneously. Notably, species that produce larger spores in the root exudate such as Glomus caledonium,79,90 Acaulospora rehmii,91 and Gigaspora margarita44,61 require a longer duration for spore initiation compared to species with smaller spores such as G. intraradices,90 G. versiforme,92 or G. proliferum.93
Root organ culture for the production of arbuscular mycorrhizal (AM) fungi provides an approach for generating a significant quantity of spores and propagules, but standardization for mass production is still needed. Various factors affecting large-scale AMF production through root organ culture have been researched and documented. In this study, the researchers achieved a significant increase in the yield of viable propagules by optimizing parameters such as gelling agent (gellan gum at 0.23% w/v), cultivation medium (M medium at pH 5.5), starter inoculum (0.13 g DW/l), humidity conditions (RH at 50%), incubation period (85-88 days), and drying temperature (25 °C). The established standardized methods proposed in this research are expected to greatly influence the manufacturing of AMF on a commercial scale. These refined techniques are anticipated to aid in the production of AMF in larger quantities.55
In vitro multiplication of hairy roots in liquid culture
The cultivation of hairy roots in liquid culture medium, such as submerged culture systems, has been the subject of numerous investigations.94,95 Large amounts of liquid medium must be used in mist conditions (involving sprayed liquid medium), and these volumes must be regularly refilled. A rotating drum containing root organs in liquid culture media that was oxygenated by vibration or spinning stir was used in investigations by Kondo et al. However, none of these methods have been used to produce AMF because the stress caused by vibration and whirling stir prevents AMF from developing.96
Carrot roots (Daucus carota) have been used to grow AMF (Rhizophagus irregularis, formerly known as “Glomus intraradices”) in a liquid culture media using an airlift bioreactor.78 However, there are several difficulties because of the system’s intricacy and the need for a steady flow of sterile air. Furthermore, it is difficult to harvest, sample, and subculture AMF-colonized root sections using this method, which makes continuous cultivation challenging. Furthermore, just 20,000 spores per litre of medium is the comparatively poor spore yield obtained using this method. The equivalent liquid culture Petri plate method yields 30,000 spores per litre, which is more than this amount.97 As a result, both methods are nearly ten times less effective than the standard ROC in split plates, which can generate over 500,000 spores per litre after four months;85 the entire yield is reduced when compared to the split plate’s total medium.
The mycorrhization rate, which ranged from 25% to 75%, did not exceed that of the same liquid culture medium in Petri plate culture or normal ROCs, and the spore germination rate, which was only 58% in the airlift bioreactor, was comparatively low. Furthermore, the spore density, hyphae distribution, and degree of root colonization varied significantly among the root material colonized by AMF. Both the liquid media and the AMF-colonized roots turned brown after 69 and 93 days, and neither the AMF nor the roots continued to develop after this point.97 This method of AMF culture has a lower probability of being used in large-scale production.
Approaches for enhancing AM fungal sporulation under in vitro conditions through the ROC technique
Effects of the addition of phenolic compounds on the production of AM fungal propagules
Plants include a large class of secondary metabolites called phenolic compounds. They have a variety of structures, ranging from simple ones like phenolic acids to polyphenols, which are flavonoids with multiple groups, and polymeric compounds, which depend on these different classes.98 These compounds include phenolic acids, lignans, quinones, coumarins, tannins, stilbenes, flavonoids, and curcuminoids.99 These substances frequently serve as AMF colonization signalling molecules.100
The phenylpropanoid family, which includes a diverse array of secondary metabolites, encompasses a large group of compounds known as flavonoids. These substances serve a variety of biological purposes in plants and serve as signalling molecules in the rhizosphere’s beneficial plant-microbe interactions.101 To date, more than ten thousand unique flavonoids have been discovered. The chemical structures of these compounds permit their subclassification into essential categories, including chalcones, anthocyanins, flavones, isoflavonoids, flavanonols, and flavanones.102
Certain essential signalling elements are common to both the presymbiotic and symbiotic stages of Rhizobium-legume and arbuscular mycorrhizal (AM) symbiosis.103,104 Initially, these components were considered to have little significance for establishing AM.105 But later studies by Akiyama et al. Scervino et al. and Steinkellner et al. showed that certain flavonoids contribute to either root colonization or spore germination. Additional research has also demonstrated that flavonoids have a role in AM symbiosis as signalling chemicals. It has been shown that these flavonoids enhance hyphal development, leading to a rise in secondary spore generation, hyphal growth, branching, and spore germination. Notably, it has been demonstrated that some flavonoid compounds enhance AM colonization in roots. Communication between fungi and plants depends on these flavonoid molecules.106-108
In Glomus etunicatum, the addition of quercetin, which is a flavonoid and antioxidant, led to improvements in hyphal elongation, hyphal branching, and spore germination. Moreover, both 4′,7-dihydroxyflavone and 4′,7-dihydroxyflavanone were found to enhance spore germination.109 In addition to promoting spore germination, flavan-3-ol facilitated the AM root colonization, mycelia development, and ramification of several AM fungi. When exposed to flavonols such as morin, luteolin, and chrysin, Gigaspora and Glomus showed further improvements in their presymbiotic hyphal development.110
The flavonoid catechin anhydrous was found to have the greatest effect on arbuscule quantity and the mycorrhizal root colonization rate. This conclusion is reinforced by findings that some flavonoids can improve spore germination, mycelial expansion, and AMF colonization of roots.111 Similarly, in Gigaspora margarita, the flavonoids apigenin, hesperetin, and naringenin promote root colonization, hyphal growth, and spore germination.112
The impacts of various phenolic compounds, including coumarin, esculetin, ferulic acid, protocatechuic acid, cinnamic acid, catechin anhydrous, tannic acid, and catechol, on the production of AM fungal propagules have been investigated. In terms of fungal propagule generation, phenolic chemicals respond differently when combined with media pH. Two pH values (5.7 and 6.5) were used in this study, which was carried out in both liquid and solid media. Depending on the phenolic compounds used, mycorrhizal frequency (F%) varied significantly, ranging from 53.33% to 100.00%. The most significant results were obtained with protocatechuic acid and catechol.53
Certain flavonoids have been shown in numerous investigations to prevent spore germination. The hyphal growth from Gi. margarita spores that germinated over a 10-42-day period were improved by quercetin (10 µM). There were 13 auxiliary cells on average per spore and more than 500 mm of hyphal growth. Quercetin also helped the growth of Glomus etunicatum. Quercetin’s glycosides, including quercitrin and rutin, lacked stimulatory properties. The axenic growth of Gi. margarita under well-regulated conditions is the largest vesicular-arbuscular mycorrhizal fungal growth ever recorded.113 Another study, however, found that the frequency of entry points and root colonization of R. irregularis, Funneliformis mosseae, and Gi. margarita was stimulated during this period.107 Since these variables are crucial when using signalling substances, the results seem to differ depending on the fungal genotype, experimental setup, and probably the amounts used. In vitro tests showed that at different dosages, flavonoids like medicarpin, quercetin chrysin, genistein, and rutin improved spore germination and hyphal growth in the AM fungus R. irregularis. The fact that chrysin only had a stimulating impact at low (‘physiological’, nanomolar range) doses suggests that it might serve as a signalling molecule produced from plants during the development of AM symbioses.114
Studies conducted in vitro on several flavonols, most notably quercetin, showed that they stimulated the hyphal development and spore germination of the AM fungus Gi. margarita.113,115 Other AM fungi, such as Gi. rosea115 and Gi. gigantea,116 as well as F. mosseae,117 Claroideo glomus etunicatum,109,113 G. macrocarpum,109 and R. irregularis, have been known to benefit from quercetin’s ability to promote spore germination and hyphal growth.113,118 At high concentrations, these effects were consistently seen.100 But Lidoy et al. showed that even at low doses, chrysin, quercetin (0.01 and 0.1 µM), and rutin (0.01 µM) might promote fungal growth. Rutin led to a 2.5-fold increase in spore germination at a concentration of 0.01 µM, whereas quercetin exhibited a comparable effect at 0.1 µM.114 Few studies have noted the absence of an effect of rutin on fungal development, which may be attributed to the use of high concentrations of the compound in those experiments.46,107,113 These results suggest that flavonoids have a favourable effect on hyphal development and spore germination; however, the concentration-dependent effect emphasises how crucial flavonoid levels are for mycorrhizal characteristics.114 In the presymbiotic phase of AM symbiosis, these flavonoids may function as signalling chemicals. This may facilitate the development of hyphal growth, spore germination, and symbiosis. As a result, the addition of flavonoids improves plant-AM interactions in vitro and may improve commercial AM fungal-based inoculants.
Addition of fatty acids stimulates the AM propagule development
Acquiring carbon sources from their host is crucial for AM fungal energy production and the formation of fungal cell components. Recent studies have revealed that AM fungi are capable of absorbing lipids from their host through arbuscles119,120; nevertheless, less has been determined about the precise mechanisms by which these fungi use these lipids as nutrition. The inability of AM fungi to produce long-chain fatty acids on their own is suggested by the absence of genes for cytosolic fatty acid synthases in their genomes, which are required for the de novo synthesis of fatty acids.121,122 On the other hand, during AM symbiosis, plants produce more fatty acids and use arbuscles to transfer lipids to AM fungus, potentially in the form of β-monoacylglycerol (β-MAG).119,120 These results demonstrate that the development of AM fungus depends on exogenous fatty acids. Numerous chemicals have already been found to be hyphal branching triggers during the presymbiotic stage, including palmitoleic acid, strigolactones, 2-hydroxy fatty acids, and branched fatty acids.
A study used a variety of fatty acids to examine the growth and spore production of AM fungus in non-symbiotic environments. Only myristate, one of the eight fatty acids that were assessed, clearly promoted widespread hyphal growth and the generation of secondary spores that are compatible with infection in an asymbiotic environment. Myristate also provided R. irregularis with carbon and energy for hyphal development. Using fatty acids, this study showed that AM fungal biomass increased in a pure culture setting. Furthermore, myristate promoted the growth of R. clarus and Gi. margarita and facilitates the asymbiotic growth of a wider variety of AM fungal species.123 The development of AM fungal host-free cultures was made easier by the use of palmitoleic acid.124 The size of the spores produced under myristate treatment is smaller than those produced symbiotically, even though myristate encourages AM fungal growth and sporulation. Similar results were observed for palmitoleic acid-induced spores. A compelling future issue for growing AM fungus in pure systems is attaining spore maturation without a host, as these smaller spores have reduced germination rates and infectivity. These spores can serve as both inocula that establish symbiotic relationships with plants and propagules for future generations.123
Effects of the addition of phytohormones on AM fungal propagule development
The interactions between arbuscular mycorrhizal (AM) fungus and their host plants are enhanced by specific phytohormones. Signalling molecules called “branching factors” (BFs) are released by the roots of host plants. These molecules were first discovered in the root exudates of Lotus japonicus and were found to be strigolactone (5-deoxy-strigol).125 Previously identified as stimulants for seed germination in the root-parasitic weeds Striga and Orobanche, strigolactones (SLs) are a class of sesquiterpene lactones.126 AM fungus need their host plants’ signalling chemicals in addition to nutrition to develop and proliferate.127-129
Apart from their function as rhizosphere signalling molecules, strigolactones are a kind of plant hormone that affects various facets of flowering plant growth and development.130,131 strigolactones are distinct from other plant hormones due to their dual function in the rhizosphere as hormones and signalling molecules. Strigolactones (SLs) act as precolonization signals from the host that prompt hyphal branching and enhance energy metabolism in AM fungi, facilitating their shift from an asymbiotic to a presymbiotic growth stage.132 Strigolactones support AM symbiosis through potential secreted proteins.133
Numerous physiological effects of strigolactones and arbuscular mycorrhizal fungi (AMF) have been documented. The presence of strigolactones stimulated spore germination in both Glomus intraradices and Glomus claroideum, whereas in G. rosea, alterations in mitochondrial shape and movement occured, resulting in increased density within the fungal cells.19 Strigolactones play a dual role, acting as key signals for AMF hyphal branching and directing their development towards plant roots. These chemicals can promote hyphal branching in AM fungi at remarkably low concentrations, demonstrating that AM fungi have a sensitive detection mechanism for strigolactones.134
During the colonization of roots by AM fungi, the levels of methyl jasmonate (MeJA) increase, along with the activation of genes responsible for jasmonic acid biosynthesis in the cortical cells of plants containing highly branched arbuscules that facilitate nutrient exchange.19 This observation has inspired research into the potential use of jasmonic acid to increase both colonization and sporulation under symbiotic conditions.
The production of asymbiotic spores (AS) in R. clarus was increased through the combination of two phytohormones, strigolactone and jasmonate, when myristate and organic nitrogen peptone were also present. Myristate and peptone were shown to promote hyphal branching and elongation in R. clarus, with peptone being particularly critical for facilitating sporulation. The use of GR24, a strigolactone analogue, alongside jasmonates has been shown to act synergistically, increasing both the quantity and size of secondary spores. Although asymbiotic spores can be carried over to future generations and can serve as inocula for establishing a symbiotic relationship with plants, they are still smaller in size than symbiotic spores are.135
Coculture with bacteria improves AM propagule development
AM fungi are thought to be incapable of being cultivated without a host. However, research has shown that AM fungi may fulfil their life cycle without host plants when cocultivated with the mycorrhiza-helper bacteria Paenibacillus validus and the AM fungus Rhizophagus irregularis.136 This cocultivation encourages the development and branching of hyphae, leading to the production of secondary infectious spores that do not depend on host plants. It has been proposed that diffusible substances produced by bacteria may induce hyphal differentiation. The branching of hyphae in R. irregularis is also enhanced when a cell-free extract from P. validus is used.124 The compound 12-methyl tetradecanoic acid 4,5, a branched-chain fatty acid sourced from bacterial cultures, fosters both the branching of hyphae from mother spores and the generation of secondary spores in an axenic culture of Rhizophagus irregularis. Raffinose, a trisaccharide present in bacterial cultures, promotes hyphal development in AM fungi but does not encourage branching or sporulation.136 These findings also provide insights into the cultivation of AM fungi without living hosts.
Autotrophic system for the production of AM fungi using superabsorbent polymer
AM fungi can be grown in a clear, sterile environment using an autotrophic culture system, which combines the advantages of in vitro and in vivo methods. During this procedure, the AM fungus infects the roots of micropropagated plantlets as the shoots emerge. Broad root colonization, a vast extraradical mycelium, and thousands of spores can all be produced by the system. Under the same circumstances, the spores were able to colonize additional plantlets. These results validate the autotrophic culture system’s capacity to sustainably cultivate arbuscular mycorrhizal fungus and could be a valuable instrument for examining numerous aspects of symbiosis that require a source-sink relationship and photosynthetically active tissues.137
The rhizosphere is where arbuscular mycorrhizal (AM) fungi reproduce and are permanently connected to plant roots, making them challenging to deal with and study. It is common practice to cultivate AM fungus in vitro on Petri dishes with Ri Transfer-DNA altered roots (heterotrophic hosts) or in vivo in pot cultures with an autotrophic host. Furthermore, pot cultures of AM fungus are cultivated in nonsterile, opaque conditions. In vitro cultivation, on the other hand, entails growing AM fungus in a transparent, sterile environment. Recent research has shown that the superabsorbent polymer-based autotrophic system (SAP-AS) combines the benefits of both strategies while avoiding their respective drawbacks (sterility, heterotrophic host, and opacity).138
The ability of a SAP-based autotrophic culture to multiply AMF in symbiosis with a whole plant was investigated. Because SAP-based autotrophic cultures are nutrient-deficient, a fertilization solution is required to encourage host growth. Thus, two modified minimum (M) solutions and reverse osmosis-filtered water were used to hydrate the SAP grains. These solutions’ effects on spore germination were assessed. Seven AMF species were grown on two-compartment Petri dishes using Plantago lanceolata as the host to illustrate the advantages of using the SAP-based autotrophic system to initiate and maintain AMF single-spore cultures.139
Current in vitro and in vivo culture techniques can be replaced with SAP-based autotrophic culture. The two-compartment Petri dishes used in this approach make it simple to start single-spore cultures and track the spread of fungi. Seven AM fungal species (Gigaspora rosea, Diversispora varaderana, Sclerocystis sp., Funneliformis geosporus, Rhizophagus irregularis, Racocetra fulgida, and R. intraradices) from six genera and three families were grown as single-spore cultures in the SAP-based autotrophic system. The cultures were kept in nonsterile conditions for several months. In addition to avoiding cross-contamination, the Petri dishes can be stacked to maximize available area. The SAP grains that had colonized with fresh spores were utilized as inocula to start new cultures in the SAP-based system. The SAP-based autotrophic culture method is a low-cost and low-tech approach, which makes the study of AMF much more accessible.139
Using strawberry plants as the host, a polymeric hydrogel was used to produce the Glomus intraradix inoculum in plastic boxes.140 Five to seven weeks following inoculation, a large amount of sporulation was seen in the hydrogel matrix. Hydrogels are superabsorbent polymers (SAPs) derived from acrylamide monomers.141 The use of SAPs and the development of autotrophic cultivation systems are highly useful for AM fungal studies. However, further research is needed for optimization.
Arbuscular mycorrhizal fungi which interact with plants and the soil microbiome, are important for agricultural sustainability and ecological health. This review highlights various AM production methods, including substrate-based and substrate-free production methods, such as aeroponics, hydroponics, and nutrient film technologies. Sporulation and the generation of viable propagules are the most important factors determining AMF inoculum production, which can be performed in in vitro or in vivo. Substrate selection, host fungal preference, ambient factor management, and hygienic conditions must all be considered when producing through substrates. In aeroponic and hydroponic systems, essential issues to consider include the composition of the nutrient solution, pH and EC, sanitary conditions, contamination management, algal growth, and the use of appropriate nozzles. In the case of ROC-based production, effective practices such as host plant selection for explants, media selection, concentration of the carbon source, type and concentration of gelling agent, suitable fungal partner, maintenance of appropriate environmental conditions, and incorporation of certain signalling agents such as phenolics, fatty acids, and phytohormones that can increase sporulation and play a significant role in determining the quality of AMF inoculum and economic viability are addressed in this work. Recently developed unique methods of production in superabsorbent autotrophic systems are also discussed. When high-quality inoculum is produced, all of these impacting elements must be considered to increase the production of AMF spores and propagules. Leveraging AMF life cycle dynamics and interactions in the field might increase agricultural resilience and productivity, increasing global food security and ecosystem health.
Constraints and suggestions for in vivo and in vitro AM production methods
In vivo production is the process of culturing AM fungi with host plants in substrate-based or soil-based systems. This technique is susceptible to contamination by other soil microbes, which may hinder or compete with AM fungi for growth. The soil, pots, and ongoing care of plants and their growing conditions are among the many resources and space requirements. On a large scale, this makes it expensive and labor intensive. Variations in host plant health, soil characteristics, and the environment can affect fungal inoculum quality and quantity.53 Under in vitro methods, AM fungi are grown on transformed roots (e.g., hairy roots generated by Agrobacterium rhizogenes) via the ROC technique. To avoid contamination, this highly intricate procedure requires sterile settings. Long-term in vitro propagation may result in domestication, a process in which fungi may change genetically and functionally, perhaps decreasing their efficacy upon reintroduction into agricultural or natural environments. High initial setup costs result from the need for specialized facilities and equipment to establish in vitro cultures. In addition, the most important aspect is the need for technical expertise as well as continuous monitoring like nurturing a child. The labour-intensive aspect of maintaining sterile cultures and the requirement for exact control over growing conditions make scaling up in vitro production to suit large-scale demands difficult.81
Despite the advantages and disadvantages of both production methods, one should follow the process of maximising production and choose production methods on the basis of the resources available. Thus, only the ultimate objective of producing high-quality inoculum been accomplished. From this perspective, the present review sheds light on the production of quality inocula in a sustainable manner.
Improved cultivation techniques and cutting-edge technology can be used in tandem to overcome the limitations in the mass production of arbuscular mycorrhizal fungi. In in vivo production, symbiotic efficiency can be improved and environmental variability can be decreased by using controlled environment agriculture and optimizing host plant selection. While automated harvesting and modular growth methods might increase scalability and decrease labour intensity, sterilized substrates and routine monitoring can reduce the danger of infection. Improved medium formulations and genetic optimization can increase fungal growth and lessen the impacts of domestication in in vitro production. Cost-effective materials and bioreactor technology can be used to increase production while lowering expenses. The use of a variety of fungal strains and gradual acclimatization can improve adaptability and functional diversity, while strict sterile procedures and regular screening can guarantee culture integrity. Together, these tactics solve the biological, economic, and technical obstacles to AM fungal mass production, increasing its sustainability and efficiency.
AM fungi (AMF) are known for their obligatory nature which requires root colonization for the completion of their life cycle, which includes the formation of propagules, which are asexual spores. Improving nutrient absorption (especially phosphorus) and plant health plays an important role in sustainable agriculture. However, issues in production, such as contamination, confined sporulation, and dependence on host plants impede large-scale use. All the parameters discussed in this review strengthen the quality of AM inoculum while also increasing production, allowing the inoculum to be used without any hindrance.
While undertaking substrate-based production, dual or tripartite culturing of the inoculum with beneficial bacteria would improve the quality of AMF as well as the incorporation of bacterial inoculants, resulting in improved colonization upon application. The production of high-quality inoculum with the inclusion of signalling molecules may also be considered to improve production under in vitro conditions as well as efficiency and interaction with host plants during field application. Understanding and applying AMF-plant signalling networks will also enable more targeted manipulation to further increase crop productivity and nutrient uptake. Another constraint observed under in vitro production is the development of small spores compared with substrate-based as well as field-grown spores. Smaller spores exhibit less infectivity and germination, which reduces further development and multiplication of AM spores. Reduced infectivity and colonization may also affect the potential benefits of AM fungal application to crop plants. Studies are needed to compare the germination, infection potential, proliferation and efficacy of AM spores in vitro and in vivo as well as in the field. Additionally, ways to increase the size and infectivity of spores produced via in vitro culture methods need to be explored. In addition to production, formulation and delivery have a greater influence on crop productivity. Formulating suitable products unique to the system benefits crop productivity more than using a single formulation for all applications. These elements require further research to develop crop and system-specific formulations and delivery methods, which will increase the benefits of AMF in all spheres of agriculture.
ACKNOWLEDGMENTS
The authors acknowledge the support of Tamil Nadu Agricultural University, Coimbatore, Tamil Nadu for organizing the technical writing workshops to facilitate the students’ writing this review article.
CONFLICT OF INTEREST
The authors declare that there is no conflict of interest.
AUTHORS’ CONTRIBUTION
KK conceptualized the study. MP wrote the manucript. KK, RTV, RS, RA, SMPK and IYR reviewed and edited the manuscript. All authors read and approved the final manuscript for publication.
FUNDING
None.
DATA AVAILABILITY
Not applicable.
ETHICS STATEMENT
Not applicable.
- Parniske M. Arbuscular mycorrhiza: the mother of plant root endosymbioses. Nat Rev Microbiol. 2008;6(10):763-775.
Crossref - Remy W, Taylor TN, Hass H, Kerp H. Four hundred-million year-old vesicular arbuscular mycorrhizae. Proc Natl Acad Sci U S A. 1994;91(25):11841-11843.
Crossref - Simon L, Bousquet J, Levesque RC, Lalonde M. Origin and diversification of endomycorrhizal fungi and coincidence with vascular land plants. Nature. 1993;363:67-59.
Crossref - Bago B, Pfeffer PE, Shachar-Hill Y. Carbon metabolism and transport in arbuscular mycorrhizas. Plant Physiol. 2000;124(3):949-958.
Crossref - Ho-Plagaro T, Garcia-Garrido JM. Molecular regulation of arbuscular mycorrhizal symbiosis. Int J Mol Sci. 2022;23(11):5960.
Crossref - Hung LL, Sylvia DM. Production of vesicular-arbuscular mycorrhizal fungus inoculum in aeroponic culture. Appl Environ Microbiol. 1988;54(2):353-357.
Crossref - Sylvia DM, Hubbell DH. Growth and sporulation of vesicular-arbuscular mycorrhizal fungi in aeroponic and membrane systems. Symbiosis. 1986;1:259-267.
- Strullu DG, Plenchette C. Encapsulation of the intraradical form of Glomus in alginate and use of the capsules as inoculum. C R Acad Sci Paris. 1990; 310:447-452.
- Mosse B, Hepper C. Vesicular arbuscular mycorrhizal infections in root organ cultures. Physiol Plant Pathol. 1975;5(3):215-223.
Crossref - Strullu GD, Romand C. Methods for obtaining endomycorrhizae with vesicles and arbuscules under axenic conditions. C R Acad Sci Paris. 1986; 303:245-250
- Strullu GD, Romand C. Axenic culture of isolated vesicles from endomycorrhizae and in vitro reassociation with tomato roots. C R Acad Sci Paris. 1987; 305:15-19.
- Becard G, Fortin JA. Early events of vesicular arbuscular mycorrhizal formation on Ri T-DNA transformed roots. New Phytol. 1988;108(2):211-218.
Crossref - Fortin JA, Becard G, Declerck S, et al. Arbuscular mycorrhiza on root-organ cultures. Can J Bot. 2002;80(1):1-20.
Crossref - Gianinazzi-Pearson V. Plant cell responses to arbuscular mycorrhizal fungi: Getting to the roots of the symbiosis. Plant Cell. 1996;8(10):1871-1883.
Crossref - Requena N, Serrano E, Ocon A, Breuninger M. Plant signals and fungal perception during arbuscular mycorrhiza establishment. Phytochemistry. 2007;68(1):33-40.
Crossref - Giovannetti M, Sbrana C, Logi C. Early processes involved in host recognition by arbuscular mycorrhizal fungi. New Phytol. 1994;127(4):703-709.
Crossref - Giovannetti M, Sbrana C, Avio L, Citernesi AS, Logi C. Differential hyphal morphogenesis in arbuscular mycorrhizal fungi during pre infection stages. New Phytol. 1993;125(3):587-593.
Crossref - Smith SM. Q&A: What are strigolactones and why are they important to plants and soil microbes? BMC Biol. 2014;12:1-7.
Crossref - Besserer A, Puech-Pages V, Kiefer P, et al. Strigolactones stimulate arbuscular mycorrhizal fungi by activating mitochondria. PLoS Biol. 2006;4(7):e226.
Crossref - Buee M, Rossigno M, Jauneaul A, Ranjeva R, Becard G. The pre-symbiotic growth of arbuscular mycorrhizal fungi is induced by a branching factor partially purified from plant root exudates. Mol Plant Microbe Interact. 2000;13(6):693-698.
Crossref - Thirkell TJ, Pastok D, Field KJ. Carbon for nutrient exchange between arbuscular mycorrhizal fungi and wheat varies according to cultivar and changes in atmospheric carbon dioxide concentration. Glob Change Biol. 2020;26(3):1725-1738.
Crossref - Ivanov S, Harrison MJ. Accumulation of phosphoinositides in distinct regions of the periarbuscular membrane. New Phytol. 2019;221(4):2213-2227.
Crossref - Breuninger M, Requena N. Recognition events in AM symbiosis: analysis of fungal gene expression at the early appressorium stage. Fungal Genet Biol. 2004;41(8):794-804.
Crossref - Gianinazzi S, Schuepp H, Barea JM, Haselwandter K. Mycorrhizal technology in agriculture: from genes to bioproducts. Springer Science & Business Media. 2002.
Crossref - Johansson JF, Paul LR, Finlay RD. Microbial interactions in the mycorrhizosphere and their significance for sustainable agriculture. FEMS Microbiol Ecol. 2004;48(1):1-13.
Crossref - IJdo M, Cranenbrouck S, Declerck S. Methods for large-scale production of AM fungi: past, present, and future. Mycorrhiza. 2011;21(1):1-16.
Crossref - Mukhongo RW, Tumuhairwe JB, Ebanyat P, et al. Production and use of arbuscular mycorrhizal fungi inoculum in sub-Saharan Africa: challenges and ways of improving. Int J Soil Sci. 2016;11(3):108-122.
Crossref - Akhtar MS, Abdullah SNA. Mass production techniques of arbuscular mycorrhizal fungi: major advantages and disadvantages: a review. Biosci Biotechnol Res Asia. 2014;11(3):1199-1204.
Crossref - Sylvia DM and Jarstfer AG. Sheared-root inocula of vesicular-arbuscular mycorrhizal fungi. Applied and Environmental Microbiology. 1992;58(1):229-232.
- Douds DD Jr, Nagahashi G, Pfeffer PE, Kayser WM, Reider C. On-farm production and utilization of arbuscular mycorrhizal fungus inoculum. Can J Plant Sci. 2005;85(1):15-21.
Crossref - Douds DD Jr, Nagahashi G, Pfeffer PE, Reider C, Kayser WM. On-farm production of AM fungus inoculum in mixtures of compost and vermiculite. Biores Tech. 2006:97:809-818.
Crossref - Gaur A, Adholeya A. Arbuscular-mycorrhizal inoculation of five tropical fodder crops and inoculum production in marginal soil amended with organic matter. Biol Fertil Soils. 2002;35(3):214-218.
Crossref - Sylvia DM and Jarstfer AG. Production of inoculum and inoculation with arbuscular Mycorrhizal fungi, 1994 in Management of Mycorrhizas in Agriculture, Horticulture and Forestry, eds AD Robson, LK Abbott and N.Malajczuk (Netherlands: Kluwer), 231-238
- Hawkins HJ, George E. Hydroponic culture of the mycorrhizal fungus Glomus mosseae with Linum usitatissimum L., Sorghum bicolor L. and Triticum aestivum L. Plant Soil. 1997;196:143-149.
Crossref - Mosse B and Thompson JP. National Research Development Corp UK, 1981. Production of mycorrhizal fungi. U.S. Patent 4,294,037.
- Lee YJ, George E. Development of a nutrient film technique culture system for arbuscular mycorrhizal plants. HortScience. 2005;40(2):378-380.
Crossref - Zobel RW, Del Tredici P, Torrey JG. Method for growing plants aeroponically. Plant Physiol. 1976;57(3):344-346.
Crossref - Mosse B. The establishment of vesicular-arbuscular mycorrhiza under aseptic conditions. Microbiology. 1962;27(3):509-520.
Crossref - Mosse B. The regular germination of resting spores and some observations on the growth requirements of an Endogone sp. causing vesicular-arbuscular mycorrhiza. Trans Brit Mycol Soc. 1959;42(3):274-286.
Crossref - Mugnier J, Mosse B. Vesicular-arbuscular mycorrhizal infection in transformed root-inducing T-DNA roots grown axenically. 1987;77(7):1045-1050.
Crossref - Chilton MD, Tepfer DA, Petit A, David C, Casse-Delbart F, Tempe J. Agrobacterium rhizogenes inserts T-DNA into the genomes of the host plant root cells. Nature. 1982;295:432-434.
Crossref - Broothaerts W, Mitchell HJ, Weir B, et al. Gene transfer to plants by diverse species of bacteria. Nature. 2005;433(7026):629-633.
Crossref - Tepfer DA, Tempe J. Production of agropine by roots formed under the action of Agrobacterium rhizogenes strain A4. C R Acad Sci Paris. 1981;292(3);153-156.
Crossref - Diop TA, Becard G, Piche Y. Long-term in vitro culture of an endomycorrhizal fungus, Gigaspora margarita, on Ri T-DNA transformed roots of carrot. Symbiosis. 1992;12(1992):249-259.
- Declerck S, Strullu DG, Plenchette C. Monoxenic culture of the intraradical forms of Glomus sp. isolated from a tropical ecosystem: a proposed methodology for germplasm collection. Mycologia. 1998;90(4):579-585.
- Chabot S, Bel-Rhlid R, Chenevert R, Piche Y. Hyphal growth promotion in vitro of the VA mycorrhizal fungus, Gigaspora margarita Becker & Hall, by the activity of structurally specific flavonoid compounds under CO2-enriched conditions. New Phytologist. 1992;122(3):461-467.
Crossref - Pawlowska TE, Douds DD, Charvat I. In vitro propagation and life cycle of the arbuscular mycorrhizal fungus Glomus etunicatum. Mycol Res. 1999;103(12):1549-1556.
Crossref - Raman N, Sahadevan C, Srinivasan V. Growth of AM fungus in vitro root organ culture of Sorghum vulgare and Saccharum officinarum. Indian J Exp Biol 2001;39(12):1293-1298.
- Eskandari A, Danesh YR. Study on life cycle of arbuscular mycorrhizal fungus Glomus intraradices using in vitro culturing technique. J Phytol. 2010;2(6):69-75.
- Marleau J, Dalpe Y, St-Arnaud M, Hijri M. Spore development and nuclear inheritance in arbuscular mycorrhizal fungi. BMC Evol Biol. 2011;11:149.
Crossref - Ceballos I, Ruiz M, Fernandez C, Pena R, Rodriguez A, Sanders IR. The in vitro mass-produced model mycorrhizal fungus, Rhizophagus irregularis, significantly increases yields of the globally important food security crop cassava. PLoS One. 2013;8(8):e70633.
Crossref - Srinivasan M, Kumar K, Kumutha K, Marimuthu P. Establishing monoxenic culture of arbuscular mycorrhizal fungus Glomus intraradices through root organ culture. J Appl Nat Sci. 2014;6(1):290-293.
- Abd Ellatif S, Ali EAM, Senousy HH, Razik ESA. Production of arbuscular mycorrhizal fungi using in vitro root organ culture and phenolic compounds. J Pure Appl Microbiol. 2019;13(4):1985-1994.
Crossref - Garcia KGV, de Souza OFJ, de Araujo PAP, Mendes Filho PF. Can inoculation of native arbuscular mycorrhizal fungi from a mining area attenuate stress of Acacia mangium Willd. to excess manganese? J Soils Sediments. 2024;24(9):3252-3264.
Crossref - Ghorui M, Chowdhury S, Das K, Sunar K, Prakash B. Optimizing factors for large-scale production of Arbuscular Mycorrhizal Fungi consortia using root organ cultures. J Biol Methods. 2023;10(1):jbm-e99010006.
Crossref - Hoang CK, Pham NH, Le CH, et al. Impact of nitrogen fertilizer on the mycorrhizal inoculating potential and fungal community structure in the rhizosphere of medicinal plant Curcuma longa L. Geomicrobiol J. 2019;36(5):385-395.
Crossref - Kokkoris V, Banchini C, Pare L, et al. Rhizophagus irregularis, the model fungus in arbuscular mycorrhiza research, forms dimorphic spores. New Phytologist. 2024;242(4):1771-1784.
Crossref - Hammer EC, Arellano-Caicedo C, Mafla-Endara PM, et al. Hyphal exploration strategies and habitat modification of an arbuscular mycorrhizal fungus in microengineered soil chips. Fungal Ecol. 2024;67:101302.
Crossref - Zhao J, He X, Xiao D, Chen M, Cheng M, Wang Z. Impacts of lithology and slope position on arbuscular mycorrhizal fungi communities in a karst forest soil. J Fungi. 2023;9(12):1133.
Crossref - Zhao T, Wang L, Yang J. Synergistic effects of combined application of biochar and arbuscular mycorrhizal fungi on the safe production of rice in cadmium-contaminated soil. Sci Total Environ. 2024;951:175499.
Crossref - Tiwari P, Adholeya A. In vitro co-culture of two AMF isolates Gigaspora margarita and Glomus intraradices on Ri T-DNA transformed roots. FEMS Microbiol Lett. 2002;206(1):39-43.
Crossref - Tiwari P, Adholeya A. Host dependent differential spread of Glomus intraradices on various Ri T-DNA transformed roots in vitro. Mycol Prog. 2003;2(3):171-177.
- Angelard C, Tanner CJ, Fontanillas P, Niculita-Hirzel H, Masclaux F, Sanders IR. Rapid genotypic change and plasticity in arbuscular mycorrhizal fungi is caused by a host shift and enhanced by segregation. ISME J. 2014;8:284-294.
Crossref - Ehinger M, Koch AM, Sanders IR. Changes in arbuscular mycorrhizal fungal phenotypes and genotypes in response to plant species identity and phosphorus concentration. New Phytol. 2009;184(2):412-423.
Crossref - Kokkoris V, Chagnon PL, Yildirir G, et al. Host identity influences nuclear dynamics in arbuscular mycorrhizal fungi. Curr Biol. 2021;31(7):1531.e-8.e.
Crossref - Mateus ID, Masclaux FG, Aletti C, et al. Dual RNA-seq reveals large-scale non-conserved genotype – genotype-specific genetic reprogramming and molecular crosstalk in the mycorrhizal symbiosis. ISME J. 2019;13(5):1226-1238.
Crossref - Labour K, Jolicoeur M, St-Arnaud M. Arbuscular mycorrhizal responsiveness of in vitro tomato root lines is not related to growth and nutrient uptake rates. Can J Bot. 2003;81(7):645-656.
- Yusuf NA, Rahim NSM, Azhar SZA, Ghani KA, Sommano S, Khalid N. Adventitious root cultures of Boesenbergia rotunda as a source of Pinostrobin. Int J Adv Sci Eng IT. 2018;8(2):377-383.
Crossref - Douds DD. Increased spore production by Glomus intraradices in the split-plate monoxenic culture system by repeated harvest, gel replacement, and resupply of glucose to the mycorrhiza. Mycorrhiza. 2002;12(4):163-167.
Crossref - Douds Jr DD. A procedure for the establishment of Glomus mosseae in dual culture with Ri T-DNA-transformed carrot roots. Mycorrhiza. 1997;7(2):57-61.
- Diop TA. Ecology of vesicular-arbuscular mycorrhizal fungi associated with Acacia albida (Del) in the Sahelian and Sudanian-Guinean zones of Senegal [Doctoral thesis]. Angers, France: University of Angers;1995
- Prabagaran S, Kumutha K. Standardization of parameters for root organ culturing of AM fungi [M.Sc. (Ag) Thesis]. Tamil Nadu Agricultural University, Tamil Nadu; 2008.
- Butcher DN. The culture of isolated roots. 1980 – In Tissue culture methods for plant pathologists (ed. D.S. Ingram & J.P. Helgeson), pp. 113-117. Oxford, U.K.: Blackwell.
- Becard G, Piche Y. Establishment of Vesicular-arbuscular Mycorrhiza in Root Organ Culture: Review and Proposed Methodology. Methods in Microbiology. 1992;24:89-108.
Crossref - Haritha G. In vitro association of Ri T-DNA transformed carrot roots with arbuscular mycorrhizal fungi: Effect of exogenous addition of phytohormones and secondary metabolites [Doctoral Thesis]. Division of Microbiology, ICAR-Indian Agricultural Research Institute, New Delhi; 2022.
- Hussein S, Ling APK, Ng TH, Ibrahim R, Paek KY. Adventitious roots induction of recalcitrant tropical woody plant, Eurycoma longifolia. Romanian Biotechnol Lett. 2012;17(1):7026-7035.
- D’Souza J, Rodrigues KM, Rodrigues BF. Modified Strullu and Romand (MSR) medium devoid of sucrose promotes higher in vitro germination in Rhizophagus irregularis. 2013.
- Jolicoeur M, Williams RD, Chavarie C, Fortin JA, Archambault J. Production of Glomus intraradices propagules, an arbuscular mycorrhizal fungus, in an airlift bioreactor. Biotechnol Bioeng. 1999;63(2):224-232.
Crossref - Karandashov V, Kurozovkina I, Hawkin HJ, George E. Growth and sporulation of the arbuscular mycorrhizal fungus G. caledonium in dual culture with transformed carrot roots. Mycorrhiza. 2000;10(1):23-28.
Crossref - Vilarino A, Frey B, Shuepp H. MES [2-(N-morpholine)-ethane sulphonic acid] buffer promotes the growth of external hyphae of the arbuscular mycorrhizal fungus Glomus intraradices in an alkaline sand. Biol Fert Soils. 1997;25:79-81.
- Kokkoris V, Hart M. In vitro propagation of arbuscular mycorrhizal fungi may drive fungal evolution. Front Microbiol. 2019;10:2420.
Crossref - Sahoo M, Kullu B. Optimization of in vitro root organ culture and co-culture of AM fungi Claroideoglomus claroideum. Plant Sci Res. 2020;42(1&2):10-20.
- Furlan V, Fortin JA. Effects of light intensity on the formation of vesicular arbuscular endomycorrhizas on Allium cepa by Gigaspora calospora. New Phytol. 1977;79(2):335-340.
Crossref - Hayman DS. Plant growth responses to Vesiculararbuscular mycorrhiza: vi. Effect of light and temperature. New Phytol. 1974;73(1):71-80.
Crossref - St-Arnaud M, Hamel C, Vimard B, Caron M, Fortin J. Enhanced hyphal growth and spore production of the arbuscular mycorrhizal fungus Glomus intraradices in an in vitro system in the absence of host roots. Mycol Res. 1996;100(3):328-332.
Crossref - Goh D, Martin JGA, Banchini C, MacLean AM, Stefani F. RocTest: A standardized method to assess the performance of root organ cultures in the propagation of arbuscular mycorrhizal fungi. Front Microbiol. 2022;13:937912.
Crossref - Rosikiewicz P, Bonvin J, Sanders IR. Cost-efficient production of in vitro Rhizophagus irregularis. Mycorrhiza. 2017;27:477-486.
Crossref - Mosse B. Some studies relating to “independent” growth of vesicular-arbuscular endophytes. Can J Bot. 1988;66(12):2533-2540.
Crossref - Devikrishna S, Kumutha K, Santhanakrishnan P, Priya LS. Influence of surface sterilization and cold treatment on germination of AM spores. Asian J Bio Sci. 2010;5(1):58-62.
- Declerck S, D’or D, Cranenbrouck S, Boulenge LE. Modelling the sporulation dynamics of arbuscular mycorrhizal fungi in monoxenic culture. Mycorrhiza. 2001;11(5):225-230.
Crossref - Dalpe Y, Declerck S. Development of Acaulospora rehmii spore and hyphal swellings under root-organ culture. Mycologia. 2002;94(5):850-855.
Crossref - Declerck S, Strullu DG, Plenchette C. In vitro mass-production of the arbuscular mycorrhizal fungus, Glomus versiforme, associated with Ri T-DNA transformed carrot roots. Mycol Res. 1996;100(10):1237-1242.
Crossref - Declerck S, Cranenbrouck S, Dalpe Y, et al. Glomus proliferum sp. nov.: a description based on morphological, biochemical, molecular and monoxenic cultivation data. Mycologia. 2000;92(6):1178-1187.
Crossref - McKelvey SA, Gehrig JA, Hollar KA, Curtis WR. Growth of plant root cultures in liquid and gas dispersed reactor environments. Biotechnol Prog. 1993;9(3):317-322.
Crossref - Nuutila AM, Vestberg M, Kauppinen V. Infection of hairy roots of strawberry (Fragaria x Ananassa Duch.) with arbuscular mycorrhizal fungus. Plant Cell Rep. 1995;14:505-509.
Crossref - Kondo O, Honda H, Taya M, Kobayashi T. Comparison of growth properties of carrot hairy root in various bioreactors. Appl Microbiol Biotechnol. 1989;32:291-294.
Crossref - Schuessler A. Symplanta & CO KG GmbH. System and methods for continuous propagation and mass production of arbuscular mycorrhizal fungi in liquid culture. 2016; U.S. Patent Application 14/913,909
- Cheynier V. Phenolic compounds: from plants to foods. Phytochem Rev. 2012;11(2):153-177.
Crossref - Cosme P, Rodriguez AB, Espino J, Garrido M. Plant phenolics: Bioavailability as a key determinant of their potential health-promoting applications. Antioxidants. 2020;9(12):1263.
Crossref - Vierheilig H, Bago B, Albrecht C, Poulin MP, Piche Y. Flavonoids and Arbuscular-Mycorrhizal Fungi. In: Manthey, J.A., Buslig, B.S. (eds) Flavonoids in the Living System. Advances in Experimental Medicine and Biology, vol 439. Springer, Boston, MA 1998:439:9-33.
Crossref - Hassan S, Mathesius U. The role of flavonoids in root-rhizosphere signalling: opportunities and challenges for improving plant-microbe interactions. J Exp Bot. 2012;63(9):3429-3444.
Crossref - Panche AN, Diwan AD, Chandra SR. Flavonoids: an overview. J Nutr Sci. 2016;5:e47.
Crossref - de Bruijn FJ. The common symbiotic signaling pathway (CSSP or SYM). Model Legume Medicago Truncatula. 2019:521-521.
Crossref - Mukherjee A, Ane JM. Germinating spore exudates from arbuscular mycorrhizal fungi: molecular and developmental responses in plants and their regulation by ethylene. Mol Plant Microbe Interact. 2011;24:260-270.
Crossref - Becard G, Taylor LP, Douds DD, Pfeffer PE, Doner LW. Flavonoids are not necessary plant signal compounds in arbuscular mycorrhizal symbioses. Mol Plant-Microbe Interact. 1995;8:252-258.
Crossref - Akiyama K, Matsuoka H, Hayashi H. Isolation and identification of a phosphate deficiency-induced C-glycosylflavonoid that stimulates arbuscular mycorrhiza formation in melon roots. Mol Plant Microbe Interact. 2002;15:334-340.
Crossref - Scervino JM, Ponce MA, Erra-Bassells R, et al. The effect of flavones and flavonols on colonization of tomato plants by arbuscular mycorrhizal fungi of the genera Gigaspora and Glomus. Can J Microbiol. 2007;53:702-709.
Crossref - Steinkellner S, Lendzemo V, Langer I, et al. Flavonoids and strigolactones in root exudates as signals in symbiotic and pathogenic plant-fungus interactions. Molecules. 2007;12(7):1290-1306.
Crossref - Tsai SM, Phillips DA. Flavonoids released naturally from alfalfa promote development of symbiotic Glomus spores in vitro. Appl Environ Microbiol. 1991;57(5):1485-1488.
Crossref - Aikawa J, Ishii T, Kuramoto M, Kadoya K. Growth stimulants for vesicular-arbuscular mycorrhizal fungi in satsuma mandarin pomace. J Japan Soc Hort Sci. 2000;69(4):385-389.
Crossref - Soares ACF, Martins MA, Mathias L, Freitas MSM. Arbuscular mycorrhizal fungi and the occurrence of flavonoids in roots of passion fruit seedlings. Scientia Agricola. 2005;62(4):331-336.
Crossref - Nakabayashi R, Mori T, Saito K. Alternation of flavonoid accumulation under drought stress in Arabidopsis thaliana. Plant Signal Behav. 2014;9(8):e29518.
Crossref - Becard G, Douds DD, Pfeffer PE. Extensive in vitro hyphal growth of vesicular-arbuscular mycorrhizal fungi in the presence of CO2 and flavonols. Appl Environ Microbiol. 1992;58(3):821-825.
Crossref - Lidoy J, Berrio E, Garcia M, Espana-Luque L, Pozo MJ, Lopez-Raez JA. Flavonoids promote Rhizophagus irregularis spore germination and tomato root colonization: A target for sustainable agriculture. Front Plant Sci. 2023;13:1094194.
Crossref - Scervino JM, Ponce MA, Erra-Bassells R, Vierheilig H, Ocampo JA, Godeas A. Flavonoids exhibit fungal species and genus specific effects on the presymbiotic growth of Gigaspora and Glomus. Mycol Res. 2005;109(7):789-794.
Crossref - Baptista MJ, Siqueira JO. Effect of flavonoids on the germination and asymbiotic growth of vesicular-arbuscular mycorrhizal fungi. Braz J Plant Physiol. 1997; 6(2): 127-134.
- Kape R, Wex K, Parniske M, Gorge E, Wetzel A, Werner D. Legume root metabolites and VA-mycorrhiza development. J Plant Physiol. 1993;141(1):54-60.
Crossref - Poulin MJ, Simard J, Catford JG, Librie F, Piche Y. Response of symbiotic endomycorrhizal fungi to estrogens and antiestrogens. Mol Plant-Microbe Interact. 1997;10:481-487.
Crossref - Brands M, Wewer V, Keymer A, Gutjahr C, Dormann P. The Lotus japonicus acyl acyl carrier protein thioesterase FatM is required for mycorrhiza formation and lipid accumulation of Rhizophagus irregularis. Plant J. 2018;95(2):219-232.
Crossref - Bravo A, Brands M, Wewer V, Dormann P, Harrison MJ. Arbuscular mycorrhiza-specific enzymes FatM and RAM2 fine-tune lipid biosynthesis to promote development of arbuscular mycorrhiza. New Phytol. 2017;214(4):1631-1645.
Crossref - Sun X, Chen W, Ivanov S, et al. Genome and evolution of the arbuscular mycorrhizal fungus Diversispora epigaea (formerly Glomus versiforme) and its bacterial endosymbionts. New Phytol. 2019;221(3):1556-1573.
Crossref - Wewer V, Brands M, Dormann P. Fatty acid synthesis and lipid metabolism in the obligate biotrophic fungus Rhizophagus irregularis during mycorrhization of Lotus japonicus. Plant J. 2014;79(3):398-412.
Crossref - Sugiura Y, Akiyama R, Tanaka S, et al. Myristate can be used as a carbon and energy source for the asymbiotic growth of arbuscular mycorrhizal fungi. Proc Natl Acad Sci U S A. 2020;117(41):25779-25788.
Crossref - Kameoka H, Tsutsui I, Saito K, et al. Stimulation of asymbiotic sporulation in arbuscular mycorrhizal fungi by fatty acids. Nat Microbiol. 2019;4:1654-1660.
Crossref - Akiyama K, Matsuzaki KI, Hayashi H. Plant sesquiterpenes induce hyphal branching in arbuscular mycorrhizal fungi. Nature. 2005;435(7043):824-827.
Crossref - Bouwmeester HJ, Matusova R, Zhongkui S, Beale MH. Secondary metabolite signalling in host-parasitic plant interactions. Curr Opin Plant Biol. 2003;6(4):358-364.
Crossref - Bucking H, Abubaker J, Goninddarajulu M, et al. Root exudates stimulate the uptake and metabolism of organic carbon in germinating spores of Glomus intraradices. New Phytol. 2008;180(3):684-695.
Crossref - Gianinazzi-Pearson V, Sejalon-Delmas N, Genre A, Jeandroz S, Bonfante P. Plants and arbuscular mycorrhizal fungi: cues and communication in the early steps of symbiotic interactions. Adv Bot Res. 2007;46:181-219.
Crossref - Nagata M, Yamamoto N, Miyamoto T, et al. Enhanced hyphal growth of arbuscular mycorrhizae by root exudates derived from high R/FR treated Lotus japonicus. Plant Signal Behav. 2016;11(6):e1187356.
Crossref - Umehara M, Hanada A, Yoshida S, et al. Inhibition of shoot branching by new terpenoid plant hormones. Nature. 2008;455(7210):195-200.
Crossref - Gomez-Roldan V, Fermas S, Brewer PB, et al. Strigolactone inhibition of shoot branching. Nature. 2008;455:189-194.
Crossref - Besserer A, Becard G, Jauneau A, Roux C, Sejalon-Delmas N. GR24, a synthetic analog of strigolactones, stimulates the mitosis and growth of the arbuscular mycorrhizal fungus Gigaspora rosea by boosting its energy metabolism. Plant Physiol. 2008;148(1):402-413.
Crossref - Tsuzuki S, Handa Y, Takeda N, Kawaguchi M. Strigolactone-induced putative secreted protein 1 is required for the establishment of symbiosis by the arbuscular mycorrhizal fungus Rhizophagus irregularis. Mol Plant Microbe Interact. 2016;29(4):277-286.
Crossref - Akiyama K, Hayashi H. Strigolactones: chemical signals for fungal symbionts and parasitic weeds in plant roots. Ann Bot. 2006;97(6):925-931.
Crossref - Tanaka S, Hashimoto K, Kobayashi Y, et al. Asymbiotic mass production of the arbuscular mycorrhizal fungus Rhizophagus clarus. Commun Biol. 2022;5(1):43.
Crossref - Hildebrandt U, Ouziad F, Marner FJ, Bothe H. The bacterium Paenibacillus validus stimulates growth of the arbuscular mycorrhizal fungus Glomus intraradices up to the formation of fertile spores. FEMS Microbiol Lett. 2006;254(2):258-267.
Crossref - Voets L, Dupre de Boulois H, Renard L, Strullu DG, Declerck S. Development of an autotrophic culture system for the in vitro mycorrhization of potato plantlets. FEMS Microbiol Lett. 2005;248(1):111-118.
Crossref - Pare L, Banchini C, Stefani F. Inoculating and Observing Arbuscular Mycorrhizal Cultures on Superabsorbent Polymer-Based Autotrophic Systems. JoVE. 2024;208:e66848.
Crossref - Pare L, Banchini C, Hamel C, Bernier L, Stefani F. A simple and low-cost technique to initiate single-spore cultures of arbuscular mycorrhizal fungi using a superabsorbent polymer. Symbiosis. 2022;88(1):61-73.
Crossref - Vestberg M, Uosukainen M. A new method for producing VA-mycorrhiza inoculum in a soil-free substrate. Mycologia. 1992;6(1):38.
Crossref - Braun O, Coquery C, Kieffer J, et al. Spotlight on the Life Cycle of Acrylamide-Based Polymers Supporting Reductions in Environmental Footprint: Review and Recent Advances. Molecules. 2021;27(1):42.
Crossref
© The Author(s) 2025. Open Access. This article is distributed under the terms of the Creative Commons Attribution 4.0 International License which permits unrestricted use, sharing, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.